Молекулы белка представляют собой. Строение и функции белка или чего не знала анжелина джоли
Белок каталаза выполняет в клетке функцию;
Класс
1. Наиболее распространенными в клетках живых организмов элементами являются:
2. Азот как элемент входит в состав:
3. Водород как элемент входит в состав:
4. На каком уровне организации не наблюдается различие между органическим и неорганическим миром?
5.Воды содержится больше в клетках:
6. Вода - основа жизни:
7. Вещества, хорошо растворимые в воде, называются:
8. К гидрофобным соединениям клетки относятся:
9. К углеводам моносахаридам относятся:
10. К углеводам полисахаридам относятся:
11. Основные функции жиров в клетке:
12.Белки - это биополимеры мономерами, которого являются:
13. Аминокислоты различаются:
14. В состав молекул белков входят:
15. Структура молекулы белка, которую определяет последовательность аминокислотных остатков:
16. Вторичная структура белка связана с:
17. Между первым и вторым понятием в задании существует определенная связь.. Найдите это слово Клетка: хлоропласт = растение: _______________
18. Наименее прочными структурными белка является:
20. При неполной денатурации белка первой разрушается структура:
21. Мономерами молекул ДНК являются:
22. Нуклеотиды ДНК состоят из:
23. Состав нуклеотидов ДНК отличается друг от друга содержанием:
24. Нуклеотиды ДНК содержат азотистые основания:
25. Нуклеотиды РНК состоят из:
26.Молекулы, при окислении которых освобождается много энергии:
27. Наиболее распространенными в клетках живых организмах элементами являются:
28. Углерод как элемент входит в состав:
29. Функции воды в клетке:
30. К углеводам моносахаридам относятся:
31. К углеводам полисахаридам относятся:
32. В состав молекулы ДНК входят остатки:
33. Продуктами реакции взаимодействия глицерина и высших жирных кислот являются:
34. Жиры и масла по отношению к воде обладают свойствами:
35. Белки - это:
36. В водных растворах аминокислоты проявляют свойства:
37. Первичная структура белка определяется:
38. Первичная структура белка поддерживается связями:
39. Ферменты выполняют следующие функции:
40. Биологическую активность белка определяет структура:
41. Молекулы, которые наиболее легко расщепляются в клетке с освобождением энергии:
42.Мономерами молекул нуклеиновых кислот являются:
43.Нуклеотиды молекулы ДНК содержат азотистые основания:
44. Углеводов содержится больше:
45.Нуклеотиды молекулы РНК содержат азотистые основания:
46. Жиры растворимы:
47. Молекула вещества, состоящая из нуклеотидов и имеющая вид одноцепочной нити:
48. Наиболее крупные размеры среди нуклеиновых кислот имеют молекулы:
49. Соли К важны для организма, так как:
50. Наука, изучающая функционирование организмов, называется:
51. Способность к хемоавтотрофному способу питания характерна для:
52. Вещества, служащие универсальными биологическими аккумуляторами энергии в клетке:
53. В молекуле ДНК количество нуклеотидов с цитозином составляет 15% от общего числа. Какой процент нуклеотидов с аденином в этой молекуле?
54. Аминокислотный остаток белка кодируется:
55. Последовательность нуклеотидов в одной из комплементарных цепей ДНК – АГА. Какова соответствующая ей последовательность нуклеотидов в другой цепи?:
56. В клетках грибов, как и в клетках животных, отсутствуют:
57. Органоиды клетки, отвечающие за ее передвижение:
58. Собственную ДНК имеют:
59. Из предложенных ответов выберите одно из положений клеточной теории :
61. АТФ считают основным источником энергии в клетки, так как:
62. Обмен веществ происходит в каждой живой клетке и представляет собой:
63. Что служит главным источником энергии, обеспечивающим круговорот веществ в экосистемах?
Структура белковой молекулы.
Органические вещества. Белки
Органические вещества.
О рганические соединения составляют в среднем 20–30% массы клетки живого организма. К ним относятся биологические полимеры – белки, нуклеиновые кислоты и полисахариды, а также жиры и ряд низкомолекулярных органических веществ – аминокислоты, простые сахара, нуклеотиды и т.д. Полимеры – сложные разветвленные или линейные молекулы, при гидролизе распадающиеся до мономеров . Если полимер состоит из одного вида мономеров, то такой полимер называют гомополимером , если в состав полимерной молекулы входят различные мономеры – то это гетерополимер . Если группа различных мономеров в полимерной молекуле повторяется (А,Б,В,А,Б,В,А,Б,В) – это регулярный гетерополимер , если нет повторения определенной группы мономеров – гетерополимер нерегулярный .
| |
Белки.
Из органических веществ клетки по количеству и значению на первом месте стоят белки. Белки , или протеины (от греч. протос – первый, главный) – высокомолекулярные гетерополимеры, органические вещества и распадающиеся при гидролизе до аминокислот . Значение белков настолько велико, что информация о белках включена в два наиболее популярных определения Жизни: «Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь, что приводит к разложению белка» (Ф.Энгельс). «Живые тела, существующие на Земле, представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, построенными из биополимеров – белков и нуклеиновых кислот» (М.В.Волькенштейн).
В состав простых белков (состоящих только из аминокислот) входят углерод, водород, азот, кислород и сера . Часть белков (сложные белки) образует комплексы с другими молекулами, содержащими фосфор, железо, цинк и медь – это сложные белки, содержащие помимо аминокислот еще и небелковую - простетическую группу . Она может быть представлена ионами металлов (металлопротеины - гемоглобин), углеводами (гликопротеины ), липидами (липопротеины ), нуклеиновыми кислотами (нуклеопротеины ).
Белки обладают огромной молекулярной массой : Один из белков – глобулин молока – имеет молекулярную массу 42000. Его формула С 1864 Н 3012 О 576 N 468 S 21 . Существуют белки, молекулярная масса которых в 10 и даже в 100 раз больше. Для сравнения: молекулярная масса спирта – 46, уксусной кислоты – 60, бензола – 78.
Белки представляют собой нерегулярные гетерополимеры , мономерами которых являются α-аминокислоты . В клетках и тканях обнаружено свыше 170 различных аминокислот, но в состав белков входит лишь 20 α-аминокислот . В зависимости от того, могут ли аминокислоты синтезироваться в организме, различают: заменимые аминокислоты – десять аминокислот, синтезируемых в организме и незаменимые аминокислоты – аминокислоты, которые в организме не синтезируются. Незаменимые аминокислоты должны поступать в организм вместе с пищей.
В зависимости от аминокислотного состава, белки бывают полноценными , если содержат весь набор незаменимых аминокислот и неполноценными , если какие-то незаменимые аминокислоты в их составе отсутствуют.
Общая формула аминокислот приведена на рисунке. Все α -аминокислоты при α -атоме углерода содержат атом водорода, карбоксильную группу (-СООН) и аминогруппу (-NH 2). Остальная часть молекулы представлена радикалом.
Аминогруппа легко присоединяет ион водорода, т.е. проявляет основные свойства. Карбоксильная группа легко отдает ион водорода – проявляет свойства кислоты. Аминокислоты являются амфотерными соединениями, так как в растворе они могут выступать как в роли кислот, так и оснований. В водных растворах аминокислоты существуют в разных ионных формах. Это зависит от рН раствора и от того, какая аминокислота: нейтральная, кислая или основная.
|
 Рис. 256. Образование дипептида.
Рис. 256. Образование дипептида.
Пептиды – органические вещества, состоящие из небольшого количества остатков аминокислот, соединенных пептидной связью. Образование пептидов происходит в результате реакции конденсации аминокислот (рис.).
При взаимодействии аминогруппы одной аминокислоты с карбоксильной группой другой, между ними возникает ковалентная азот-углеродная связь, которую называют пептидной . В зависимости от количества аминокислотных остатков, входящих в состав пептида, различают дипептиды, трипептиды, тетрапептиды и т.д. Образование пептидной связи может повторяться многократно. Это приводит к образованию полипептидов . Если полипептид состоит из большого количества остатков аминокислот, то его уже называют белком. На одном конце молекулы находится свободная аминогруппа (его называют N-концом), а на другом – свободная карбоксильная группа (его называют С-концом).
Структура белковой молекулы.
Выполнение белками определенных специфических функций зависит от пространственной конфигурации их молекул, кроме того, клетке энергетически невыгодно держать белки в развернутой форме, в виде цепочки, поэтому полипептидные цепи подвергаются укладке, приобретая определенную трехмерную структуру, или конформацию . Выделяют 4 уровня пространственной организации белков.
Первичная структура белка – последовательность расположения аминокислотных остатков в полипептидной цепи, составляющей молекулу белка. Связь между аминокислотами – пептидная.
|

В организме человека обнаружено порядка десяти тысяч различных белков, которые отличаются как друг от друга, так и от белков других организмов.
Именно первичная структура белковой молекулы определяет свойства молекул белка и ее пространственную конфигурацию. Замена всего лишь одной аминокислоты на другую в полипептидной цепочке приводит к изменению свойств и функций белка. Например, замена в b-субъединице гемоглобина шестой глутаминовой аминокислоты на валин приводит к тому, что молекула гемоглобина в целом не может выполнять свою основную функцию – транспорт кислорода (в таких случаях у человека развивается заболевание – серповидноклеточная анемия).
Первым белком, у которого была выявлена аминокислотная последовательность, стал гормон инсулин. Исследования проводились в Кембриджском университете Ф.Сэнгером с 1944 по 1954 год. Было выявлено, что молекула инсулина состоит из двух полипептидных цепей (21 и 30 аминокислотных остатков), удерживаемых около друг друга дисульфидными мостиками. За свой кропотливый труд Ф.Сэнгер был удостоен Нобелевской премии.
Вторичная структура – упорядоченное свертывание полипептидной цепи в α-спираль (имеет вид растянутой пружины) и β-структра (складчатый слой) . В α- спирали NH-группа данного остатка аминокислоты взаимодействует с СО-группой четвертого от нее остатка. Практически все «СО-» и «NН-группы» принимают участие в образовании водородных связей. Они слабее пептидных, но, повторяясь многократно, придают данной конфигурации устойчивость и жесткость. На уровне вторичной структуры существуют белки: фиброин (шелк, паутина), кератин (волосы, ногти), коллаген (сухожилия).Cкладчатый слой (синоним β-структра) формируется в том случае, если сегменты полипептидных цепей располагаются параллельно или антипараллельно друг другу в один слой. При этом образуется фигура, напоминающая лист, сложенный гармошкой.
Третичная структура - укладка полипептидных цепей в глобулы , возникающей в результате возникновения химических связей (водородных, ионных, дисульфидных) и установления гидрофобных взаимодействий между радикалами аминокислотных остатков. Основную роль в образовании третичной структуры играют гидрофильно-гидрофобные взаимодействия. В водных растворах гидрофобные радикалы стремятся спрятаться от воды, группируясь внутри глобулы, в то время как гидрофильные радикалы в результате гидратации (взаимодействия с диполями воды) стремятся оказаться на поверхности молекулы.У некоторых белков третичная структура стабилизируется дисульфидными ковалентными связями, возникающими между атомами серы двух остатков цистеина. На уровне третичной структуры существуют ферменты, антитела, некоторые гормоны. По форме молекулы различают белки глобулярные и фибриллярные. Если фибриллярные белки выполняют в основном опорные функции, то глобулярные белки растворимы и выполняют множество функций в цитоплазме клеток или во внутренней среде организма.
Четвертичная структура характерна для сложных белков, молекулы которых образованы двумя и более глобулами. Субъединицы удерживаются в молекуле исключительно при помощи нековалентных связей, в первую очередь водородных и гидрофобных.Наиболее изученным белком, имеющим четвертичную структуру, является гемоглобин. Он образован двумя a-субъединицами (141 аминокислотный остаток) и двумя b-субъединицами (146 аминокислотных остатков).С каждой субъединицей связана молекула гема, содержащая железо.
Многие белки с четвертичной структурой занимают промежуточное положение между молекулами и клеточными органеллами – например микротрубочки цитоскелета состоят из белка тубулина , состоящего из двух субъединиц. Трубочка удлиняется в результате присоединения димеров к торцу.Если по каким-либо причинам пространственная конформация белков отклоняется от нормальной, белок не может выполнять свои функции
Свойства белков.
Аминокислотный состав, структура белковой молекулы определяют его свойства. Белки сочетают в себе основные и кислотные свойства, определяемые радикалами аминокислот, чем больше кислых аминокислот в белке, тем ярче выражены его кислотные свойства. Способность отдавать и присоединять Н + определяют буферные свойства белков, один из самых мощных буферов - гемоглобин в эритроцитах, поддерживающий рН крови на постоянном уровне.
Есть белки растворимые (фибриноген), есть нерастворимые , выполняющие механические функции (фиброин, кератин, коллаген).
Есть белки активные в химическом отношении (ферменты), есть химически неактивные , устойчивые к воздействию различных условий внешней среды и крайне неустойчивые .
Внешние факторы (нагревание, ультрафиолетовое излучение, тяжелые металлы и их соли, изменения рН, радиация, обезвоживание) могут вызывать нарушение структурной организации молекулы белка. Процесс утраты трехмерной конформации, присущей данной молекуле белка без разрушения первичной структуры, называют денатурацией .Причиной денатурации является разрыв связей, стабилизирующих определенную структуру белка. Первоначально рвутся наиболее слабые связи, а при ужесточении условий и более сильные. Поэтому сначала утрачивается четвертичная, затем третичная и вторичная структуры. Изменение пространственной конфигурации приводит к изменению свойств белка и, как следствие, делает невозможным выполнение белком свойственных ему биологических функций.
Если денатурация не сопровождается разрушением первичной структуры, то она может быть обратимой , в этом случае происходит самовосстановление свойственной белку конформации. Такой денатурации подвергаются, например, рецепторные белки мембраны. Процесс восстановления структуры белка после денатурации называется ренатурацией. Если восстановление пространственной конфигурации белка невозможно, то денатурация называется необратимой . Разрушение первичной структуры белка называется деградацией .
Функции белков.
Благодаря сложности, разнообразию форм и состава, белки играют важную роль в жизнедеятельности клетки и организма в целом. Функции их разнообразны.
| Функция | Примеры и пояснения |
| 1. Строительная | Белки участвуют в образовании клеточных и внеклеточных структур: входят в состав клеточных мембран (липопротеины, гликопротеины), волос (кератин), сухожилий (коллаген) и т.д. |
| 2. Транспортная | Белок крови гемоглобин присоединяет кислород и транспортирует его от легких ко всем тканям и органам, а от них в легкие переносит углекислый газ; в состав клеточных мембран входят особые белки, которые обеспечивают активный и строго избирательный перенос некоторых веществ и ионов из клетки во внешнюю среду и обратно. |
| 3. Регуляторная | Гормоны белковой природы принимают участие в регуляции процессов обмена веществ. Например, гормон инсулин регулирует уровень глюкозы в крови, способствует синтезу гликогена, увеличивает образование жиров из углеводов. |
| 4. Защитная | В ответ на проникновение в организм чужеродных белков или микроорганизмов (антигенов) образуются особые белки - антитела, способные связывать и обезвреживать их. Фибрин, образующийся из фибриногена, способствует остановке кровотечений |
| 5. Двигательная | Сократительные белки актин и миозин обеспечивают сокращение мышц у многоклеточных животных |
| 6. Сигнальная | В поверхностную мембрану клетки встроены молекулы белков, способных изменять свою третичную структуру в ответ на действие факторов внешней среды, таким образом осуществляя прием сигналов из внешней среды и передачу команд в клетку. |
| 7. Запасающая | В организме животных белки как правило не запасаются, исключение: альбумин яиц, казеин молока. Но благодаря белкам в организме могут откладываться про запас некоторые вещества, например, при распаде гемоглобина железо не выводится из организма, а сохраняется в организме, образуя комплекс с белком ферритином. |
| 8. Энергетическая | При распаде 1 г белка до конечных продуктов выделяется 17,6 кДж. Сначала белки распадаются до аминокислот, а затем до конечных продуктов - воды, углекислого газа и аммиака. Однако в качестве источника энергии белки используются только тогда, когда другие источники (углеводы и жиры) израсходованы. |
| 9. Каталитическая | Одна из важнейших функций белков. Обеспечивается белками - ферментами, которые ускоряют биохимические реакции, происходящие в клетках. Например, рибулезобисфосфаткарбоксилаза катализирует фиксацию СО 2 при фотосинтезе. |
Ферменты илиэнзимы – особый класс белков, являющихся биологическими катализаторами. Благодаря ферментам биохимические реакции протекают с огромной скоростью. Скорость ферментативных реакций в десятки тысяч раз (а иногда и в миллионы) выше скорости реакций, идущих с участием неорганических катализаторов. Вещество, на которое оказывает свое действие фермент, называют субстратом .
Ферменты – глобулярные белки, по особенностям строения ферменты можно разделить на две группы: простые и сложные. Простые ферменты являются простыми белками, т.е. состоят только из аминокислот. Сложные ферменты являются сложными белками, т.е. в их состав помимо белковой части входит группа небелковой природы - кофактор . У некоторых ферментов в качестве кофакторов выступают витамины. В молекуле фермента выделяют особую часть, называемую активным центром. Активный центр – небольшой участок фермента (от трех до двенадцати аминокислотных остатков), именно в котором происходит связывание субстрата или субстратов с образованием фермент-субстратного комплекса. По завершении реакции фермент-субстратный комплекс распадается на фермент и продукт (продукты) реакции. Некоторые ферменты имеют (кроме активного) аллостерические центры – участки, к которым присоединяются регуляторы скорости работы фермента (аллостерические ферменты ).
Для реакций ферментативного катализа
характерны: 1) высокая эффективность, 2) строгая избирательность и направленность действия, 3) субстратная специфичность, 4) тонкая и точная регуляция.Субстратную и реакционную специфичность реакций ферментативного катализа объясняют гипотезы Э.Фишера (1890) и Д.Кошланда (1959). Э.Фишер (гипотеза «ключ-замок») предположил, что пространственные конфигурации активного центра фермента и субстрата должны точно соответствовать друг другу. Субстрат сравнивается с "ключом", фермент – с «замком».
Д.Кошланд (гипотеза «рука-перчатка») предположил, что пространственное соответствие структуры субстрата и активного центра фермента создается лишь в момент их взаимодействия друг с другом. Эту гипотезу еще называют гипотезой индуцированного соответствия.
Большинство неорганических катализаторов ускоряют химические реакции при очень высоких температурах, имеют максимальную эффективность в сильнокислой или сильнощелочной среде, при высоких давлениях, а большинство ферментов активны при температурах 35-45˚С, физиологических значениях кислотности раствора и при нормальном атмосферном давлении; скорость ферментативных реакций в десятки тысяч (а иногда и в миллионы раз) выше скорости реакций, идущих с участием неорганических катализаторов. Например, пероксид водорода без катализаторов разлагается медленно: 2Н 2 О 2 → 2Н 2 О + О 2 . В присутствии солей железа (катализатора) эта реакция идет несколько быстрее. Фермент каталаза (М=252000) за 1 сек. расщепляет 100 тыс. молекул Н 2 О 2 (М=34). Известно более 2000 различных ферментов, представленных белками с высокой молекулярной массой.
Скорость ферментативных реакций зависит от 1) температуры, 2) концентрации фермента, 3) коцентрации субстрата, 4) рН. Следует подчеркнуть, что поскольку ферменты являются белками, то их активность наиболее высока при физиологически нормальных условиях.
|
 Рис. . Зависимость скорости реакции от концентрации фермента, субстрата, рН, температуры
Рис. . Зависимость скорости реакции от концентрации фермента, субстрата, рН, температуры
При увеличении количества молекул субстрата скорость ферментативной реакции растет до тех пор, пока не произойдет насыщение активных центров фермента – если активный центр каталазы расщепляет в секунду 100 000 молекул субстрата, то при количестве молекул субстрата более 100 000 на активный центр скорость реакции не возрастет.
Увеличение концентрации фермента приводит к усилению каталитической активности, так как в единицу времени преобразованиям подвергается большее количество молекул субстрата.
Для каждого фермента существует оптимальное значение рН, при котором он проявляет максимальную активность (пепсин – 2,0, амилаза слюны – 6,8, липаза поджелудочной железы – 9,0). При более высоких или низких значениях рН активность фермента снижается. При резких сдвигах рН фермент денатурирует.
Скорость работы аллостерических ферментов регулируется веществами, присоединяющимися к аллостерическим центрам. Если эти вещества ускоряют реакцию, они называются активаторами , если тормозят – ингибиторами .Классификация ферментов . По типу катализируемых химических превращений ферменты разделены на 6 классов: 1) оксиредуктазы (перенос атомов водорода, кислорода или электронов от одного вещества к другому – дегидрогеназа), 2) трансферазы (перенос метильной, ацильной, фосфатной или аминогруппы от одного вещества к другому – трансаминаза), 3) гидролазы (реакции гидролиза, при которых из субстрата образуются два продукта – амилаза, липаза), 4) лиазы (негидролитическое присоединение к субстрату или отщепление от него группы атомов, при этом могут разрываться связи «С-С», «С-N», «С-О», «С-S» – декарбоксилаза), 5) изомеразы (внутримолекулярная перестройка – изомераза), 6) лигазы (соединение двух молекул в результате образования связей «С-С», «С-N», «С-О», «С-S» – синтетаза
Белки (синоним протеины ) - высокомолекулярные азотистые органические соединения, являющиеся полимерами аминокислот. Белки - основная и необходимая составная часть всех организмов.
Сухое вещество большинства органов и тканей человека и животных, а также большая часть микроорганизмов состоят главным образом из белков. Белковые вещества лежат в основе важнейших процессов жизнедеятельности. Так, например, процессы обмена веществ (пищеварение, дыхание, выделение и др.) обеспечиваются деятельностью ферментов (см.), являющихся по своей природе белками. К белкам относятся и сократительные структуры, лежащие в основе движения, напр, сократительный белок мышц (актомиозин), опорные ткани организма (коллаген костей, хрящей, сухожилий), покровы организма (кожа, волосы, ногти и т. п.), состоящие главным образом из коллагенов, эластинов, кератинов, а также токсины, антигены и антитела, многие гормоны и другие биологически важные вещества.
Роль белков в живом организме подчеркивается уже самим их названием «протеины» (греческий protos первый, первичный), предложенным Мульдером (G. J. Mulder, 1838), который обнаружил, что в тканях животных и растений содержатся вещества, напоминающие по своим свойствам яичный белок. Постепенно было установлено, что белки представляют собой обширный класс разнообразных веществ, построенных по одинаковому плану. Отмечая первостепенное значение белков для процессов жизнедеятельности, Энгельс определил, что жизнь есть способ существовании белковых тел, заключающийся в постоянном самообновлении химических составных частей этих тел.
Химический состав и структура белков
Белки содержат в среднем около 16% азота. При полном гидролизе белки распадаются с присоединением воды до аминокислот (см.). Молекулы белков представляют собой полимеры, которые состоят из остатков около 20 различных аминокислот, относящихся к природному L-ряду, то есть имеющих одинаковую конфигурацию альфа-углеродного атома, хотя их оптическое вращение может быть неодинаковым и не всегда направленным в одну сторону. Аминокислотный состав разных белков неодинаков и служит важнейшей характеристикой каждого белка, а также критерием его ценности в питании (см. раздел Белки в питании). Некоторые белки могут быть лишены тех или иных аминокислот. Например, белки кукурузы- зеин не содержит лизина и триптофана. Другие белки, напротив, очень богаты отдельными аминокислотами. Так, протамин лосося - сальмин содержит свыше 80% аргинина, фиброин шелка - около 40% глицина (аминокислотный состав некоторых белков представлен в табл. 1).
Таблица 1. АМИНОКИСЛОТНЫЙ СОСТАВ НЕКОТОРЫХ БЕЛКОВ (в весовых процентах аминокислот белка)
|
Аминокислоты |
Сальмин |
Инсулин быка |
Гемоглобин лошади |
Альбумин сыворотки быка |
Кератин шерсти |
Фиброин шелка |
Зеин |
|
Аланин |
1,12 |
7,40 |
6,25 |
4,14 |
29,7 |
10,52 |
|
|
Глицин |
2,95 |
5,60 |
1,82 |
6,53 |
43,6 |
||
|
Валин |
3,14 |
7,75 |
9,10 |
5,92 |
4,64 |
3,98 |
|
|
Лейцин |
13,2 |
15,40 |
12,27 |
11,3 |
0,91 |
21,1 |
|
|
Изолейцин |
1,64 |
2,77 |
2,61 |
11,3 |
|||
|
Пролин |
5,80 |
2,02 |
3,90 |
4,75 |
0,74 |
10,53 |
|
|
Фенилаланин |
8,14 |
7,70 |
6,59 |
3,65 |
3,36 |
||
|
Тирозин |
12,5 |
3,03 |
5,06 |
4,65 |
12,8 |
5,25 |
|
|
Триптофан |
1,70 |
0,68 |
|||||
|
Серин |
5,23 |
5,80 |
4,23 |
10,01 |
16,2 |
7 ,05 |
|
|
Треонин |
2,08 |
4 ,36 |
5,83 |
6,42 |
3,45 |
||
|
Цистин/2 |
12,5 |
0,45 |
5,73 |
11 ,9 |
0,83 |
||
|
Метионин |
0,81 |
2,41 |
|||||
|
Аргинин |
85,2 |
3,07 |
3,65 |
5,90 |
10,04 |
1,71 |
|
|
Гистидин |
5,21 |
8,71 |
0,36 |
1 ,32 |
|||
|
Лизин |
2,51 |
8,51 |
12,82 |
2,76 |
0,68 |
||
|
Аспарагиновая кислота |
6,80 |
10,60 |
10,91 |
2,76 |
4,61 |
||
|
Глутаминовая кислота |
18,60 |
8,50 |
16,5 |
14,1 |
2,16 |
29,6 |
При неполном (обычно ферментативном) гидролизе белков, помимо свободных аминокислот, образуется ряд веществ с относительно небольшими молекулярными весами, называемых пептидами (см.) и полипептидами. В белках и пептидах аминокислотные остатки соединены между собой так называемой пептидной (кислотно-амидной) связью, образуемой карбоксильной группой одной аминокислоты и аминогруппой другой аминокислоты:
В зависимости от числа аминокислот такие соединения называют ди-, три-, тетрапептидами и т. д., например:

Длинные пептидные цепи (полипептиды), состоящие из десятков и сотен аминокислотных остатков, образуют основу структуры белковой молекулы. Многие белки состоят из одной полипептидной цепи, в других белках имеется две или более полипептидных цепей, соединенных между собой и образующих более сложную структуру. Длинные полипептидные цепи одинакового аминокислотного состава могут давать огромное число изомеров за счет различной последовательности отдельных аминокислотных остатков (подобно тому как из 20 букв алфавита можно составить множество различных слов и их сочетаний). Поскольку различные аминокислоты могут входить в состав полипептидов в разных соотношениях, число возможных изомеров становится практически бесконечным, и для каждого индивидуального белка последовательность аминокислот в полипептидных цепях является характерной и уникальной. Эта последовательность аминокислот определяет первичную структуру белка, которая в свою очередь определяется соответствующей последовательностью дезоксирибонуклеотидов в структурных генах ДНК данного организма. К настоящему времени изучена первичная структура многих белков, главным образом белковых гормонов, ферментов и некоторых других биологически активных белков. Последовательность аминокислот определяют путем ферментативного гидролиза беков и получения так называемых пептидных карт при помощи двухмерной хроматографии (см.) и электрофореза (см.). Каждый пептид исследуется на концевые аминокислоты до и после обработки аминополипептидазой - специфическим ферментом, последовательно отщепляющим аминоконцевые (N-концевые) аминокислоты, и карбоксиполипептидазой, отщепляющей карбоксиконцевые (С-концевые) аминокислоты. Для определения N-концевых аминокислот применяют реактивы, соединяющиеся со свободной аминогруппой концевой аминокислоты. Обычно используют динитрофторбензол (1-фтор-2,4-динитробензол), дающий динитрофенильное производное с N-концевой аминокислотой, которое затем может быть идентифицировано после гидролиза и хроматографического разделения гидролизата. Наряду с динитрофторбензолом, предложенным Сангером (F. Sanger), применяется также обработка фенилизотиоцианатом по Эдману (P. Edman). При этом с концевой аминокислотой образуется фенилтиогидантоин, который легко отщепляется от полипептидной цепи и может быть идентифицирован. Для определения С-концевых аминокислот применяют нагревание пептида в уксусном ангидриде с тиоцианатом аммония. В результате конденсации получается кольцо тиогидантоина, включающее радикал концевой аминокислоты, который затем легко отщепить от пептида и установить характер С-концевой аминокислоты. Последовательность аминокислот в белке устанавливают на основании последовательности пептидов, полученных с применением разных ферментов и с учетом специфичности каждого фермента, расщепляющего белок по пептидной связи, образованной определенной аминокислотой. Таким образом, определение первичной структуры белка представляет собой весьма кропотливую и длительную работу. Нашли успешное применение различные методы прямого определения последовательности аминокислот при помощи рентгеноструктурного анализа (см.) или путем масс-спектрометрии (см.) производных пептидов, получаемых при гидролизе белка разными ферментами.
Пространственно полипептидные цепи часто образуют спиральные конфигурации, удерживаемые при помощи водородных связей и образующие вторичную структуру белка. Чаще всего встречается так называемая а-спираль, в которой на один виток приходится 3,7 аминокислотных остатков.
Отдельные аминокислотные остатки в одной и той же или в разных полипептидных цепях могут быть соединены между собой при помощи дисульфидных или эфирных связей. Так, в молекуле мономера инсулина (рис. 1) дисульфидными связями соединены между собой 6 и 11-й остатки цистеина А-цепи и 7 и 20-й остатки цистеина А-цепи соответственно с 7 и 19-м остатками цистеина В-цепи. Такие связи придают полипептидной цепи, имеющей обычно спирализованные и неспирализованные участки, определенную конформацию, называемую третичной структурой белка.

Рис. 1. Схема аминокислотной последовательности в молекуле мономера инсулина быка. Вверху- цепь А, внизу- цепь В. Жирными линиями обозначены дисульфидные связи; в кружках - сокращенные названия аминокислот.
Под четвертичной структурой белка подразумевают образование комплексов из мономерных белковых молекул. Так, например, молекула гемоглобина состоит из четырех мономеров (двух альфа-цепей и двух бета-цепей). Четвертичная структура фермента лактатдегидрогеназы представляет собой тетрамер, состоящий из 4 мономерных молекул. Эти мономеры бывают двух типов: Н, характерный для сердечной мышцы, и М, характерный для скелетных мышц. Соответственно встречается 5 разных изоферментов лактатдегидрогеназы, представляющих собой тетрамеры из разных сочетаний этих двух мономеров - НННН, НННМ, ННММ, НМММ и ММММ. Структура белка определяет его биологические свойства, и даже небольшое нарушение конформации может весьма существенно отразиться на ферментативной активности или других биологических свойствах белка. Тем не менее наиболее важное значение имеет первичная структура белка, определяемая генетически и в свою очередь часто определяющая высшие структуры данного белка. Замена даже одного аминокислотного остатка в полипептидной цепи, состоящей из сотен аминокислот, может весьма существенно изменить свойства данного белка и даже полностью лишить его биологической активности. Так, например, гемоглобин, встречающийся в эритроцитах при серповидноклеточной анемии, отличается от нормального гемоглобина А лишь заменой остатка глутаминовой кислоты в 6-м положении р-цепи на остаток валина, то есть заменой лишь одной из 287 аминокислот. Однако этой замены достаточно для того, чтобы измененный гемоглобин обладал резко нарушенной растворимостью и в значительной мере утратил свою основную функцию переноса кислорода к тканям. С другой стороны, в строго определенной структуре инсулина (рис. 1) характер аминокислотных остатков в 8, 9 и 10-м положениях цепи А (между двумя остатками цистеина), по-видимому, не имеет существенного значения, поскольку эти три остатка обладают видовой специфичностью; в инсулине быка они представлены последовательностью ала-сер-вал, у овцы - ала-гли-вал, у лошади - тре-гли-иле, а в инсулине человека, свиньи и кита - тре-сер-иле.
Физико-химические свойства
Молекулярный вес большинства белков составляет от 10-15 тысяч до 100 тысяч, однако имеются белки с молекулярным весом 5-10 тысяч и несколько миллионов. Условно полипептиды с молекулярным весом ниже 5 тысяч относят к пептидам. Большинство белковых жидкостей и тканей организма (например, белки крови, яиц и др.) растворимы в воде или в растворах солей. Белки обычно дают опалесцирующие растворы, которые ведут себя как коллоидные. Имея в своем составе много гидрофильных групп, белки легко связывают молекулы воды и находятся в тканях в гидратированном состоянии, образуя растворы или гели. Многие белки богаты гидрофобными остатками и нерастворимы в обычных растворителях белков. Такие белки (например, коллаген и эластин соединительной ткани, фиброин шелка, кератины волос и ногтей) имеют фибриллярный характер, и их молекулы вытянуты в длинные волокна. Растворимые белки обычно представлены молекулами клубкообразной, глобулярной, формы. Однако разделение белков па глобулярные и фибриллярные не абсолютно, поскольку некоторые белки (например, актин мышц) способны обратимо превращаться из глобулярной конфигурации в фибриллярную в зависимости от условий среды.
Подобно аминокислотам белки являются типичными амфотерными электролитами (см. Амфолиты), то есть меняют свой электрический заряд в зависимости от pH среды. В электрическом поле белки движутся к аноду или к катоду в зависимости от знака электрического заряда молекулы, который определяется как свойствами данного белка, так и pH среды. Это движение в электрическом поле, называемое электрофорезом, применяют для аналитического и препаративного разделения белка, как правило различающихся по своей электрофоретической подвижности. При определенном pH, называемом изоэлектрической точкой (см.), неодинаковом для разных белков, число положительных и отрицательных зарядов молекулы равно друг другу, и молекула в целом электронейтральна и не движется в электрическом поле. Это свойство белка используется для их выделения и очистки методом изоэлектрической фокусировки, заключающемся в электрофорезе белка в градиенте pH, создаваемом системой буферных растворов. При этом можно подобрать такое значение pH, при котором нужный белок выпадает в осадок (поскольку растворимость белка в изоэлектрической точке наименьшая), а большинство «загрязняющих» белков останется в растворе.
Помимо pH, растворимость белков существенно зависит от присутствии и концентрации солей в растворе. Высокие концентрации солей одновалентных катионов (чаще всего применяют сернокислый аммоний) осаждают большинство белков. Механизм такого осаждения (высаливания) заключается в связывании ионами солей воды, образующей гидратную оболочку белковых молекул. Вследствие дегидратации растворимость белков понижается и они выпадают в осадок. Таков же механизм осаждения белков спиртами и ацетоном. Осаждение белков высаливанием или органическими жидкостями, смешивающимися с водой, применяют для разделения и выделения белков с сохранением их природных (нативных) свойств. При определенных условиях осаждения белки можно получить в кристаллическом виде и хорошо очистить от других белков и небелковых примесей. Ряд процедур такого рода применяют для получения кристаллических препаратов многих ферментов или других белков. Нагревание растворов белков до высокой температуры, а также осаждение белка солями тяжелых металлов или концентрированными кислотами, особенно трихлоруксусной, сульфосалициловой, хлорной, приводит к коагуляции (свертыванию) белка и образованию нерастворимого осадка. При таких воздействиях лабильные молекулы белка денатурируют, теряют свои биологические свойства, в частности ферментативную активность, становятся нерастворимыми в исходном растворителе. При денатурации нарушается нативная конфигурация белковой молекулы, и полипептидные цепи образуют беспорядочные клубки.
При ультрацентрифугировании белки осаждаются в поле ускорения центробежной силы со скоростью, зависящей главным образом от размеров белковых частиц. Соответственно для определения молекулярных весов белков применяют определение констант седиментации в ультрацентрифуге, а также скорости диффузии белков, фильтрование их через молекулярные сита, определение электрофоретической подвижности при электрофорезе в специальных условиях и некоторые другие методы.
Методы обнаружения и определения белков
Качественные реакции на белках основаны на их физико-химических свойствах или на реакциях определенных химических групп в молекуле белка. Однако, поскольку в состав молекулы белка входит большое количество разнообразных химических группировок, реакционная способность белков очень велика и ни одна из качественных реакций на белки не является строго специфичной. Заключение о присутствии белка может быть сделано лишь на основании совокупности ряда реакций. При анализе биологических жидкостей, например мочи, где могут появляться лишь определенные белки и известно, какие вещества могут мешать реакции, бывает достаточно даже одной реакции для установления присутствия или отсутствия белков. Реакции на белки подразделяют на реакции осаждения и цветные реакции. К первым относится осаждение концентрированными кислотами, причем в клинической практике чаще всего применяют осаждение азотной кислотой. Характерной реакцией является также осаждение белков сульфо-салициловой или трихлоруксусной кислотами (последняя часто применяется не только для обнаружения белков, но и для освобождения жидкостей от белков). Присутствие белков может быть обнаружено также но свертыванию при кипячении в слабокислой среде, осаждением спиртом, ацетоном и рядом других реактивов. Из цветных реакций весьма характерна биуретовая реакция (см.) - фиолетовое окрашивание с ионами меди в щелочной среде. Эта реакция зависит от присутствия в белках пептидных связей, образующих с медью окрашенное комплексное соединение. Название биуретовой реакции происходит от продукта нагревания мочевины биурета (H 2 N-CO-NH-CO-NH 2), являющегося простейшим соединением, дающим эту реакцию. Ксантопротеиновая реакция (см.) заключается в желтом окрашивании осадка белков при воздействии концентрированной азотной кислотой. Окрашивание появляется вследствие образования продуктов нитрования ароматических аминокислот, входящих в состав белковой молекулы. Реакция Миллона дает ярко-красное окрашивание с солями ртути и азотистой кислотой в кислой среде. На практике обычно используют азотную кислоту, которая всегда содержит небольшую примесь азотистой. Реакция специфична для фенольного радикала тирозина и поэтому получается только с белков, содержащими тирозин. Реакция Адамкевича обусловлена радикалом триптофана. Она дает фиолетовое окрашивание в концентрированной серной кислоте с уксусной к-той (см. Адамкевича реакция). Реакция получается при замене уксусной кислоты на различные альдегиды. При использовании уксусной кислоты реакция обусловлена глиоксиловой кислотой, содержащейся в уксусной как примесь. Количественно белки определяют обычно по белковому азоту, то есть по содержанию общего азота в осадке белков, отмытом от низкомолекулярных веществ, растворимых в осадителе. Азот в биохимических исследованиях и при клинических анализах обычно определяют методом Кьельдаля (см. Кьельдаля метод). Общее содержание белка в жидкостях часто определяют колориметрическими методами, в основе которых лежат разные модификации биуретовой реакции. Часто пользуются методом Лаури, в котором применяется реактив Фолина на тирозин в сочетании с биуретовой реакцией (см. Лаури метод).
Классификация белков
Из-за относительно больших размеров белковых молекул, сложности их строения и отсутствия достаточно точных данных о структуре большинства белков еще нет рациональной химической классификации белков. Существующая классификация в значительной мере условна и построена главным образом на основании физико-химических свойств белков, источников их получения, биологической активности и других, нередко случайных, признаков. Так, по физико-химическим свойствам белки делят на фибриллярные и глобулярные, на гидрофильные (растворимые) и гидрофобные (нерастворимые) и т. п. По источнику получения белки подразделяют на животные, растительные и бактериальные; на белки мышечные, нервной ткани, кровяной сыворотки и т. п.; по биологической активности - на белки-ферменты. белки-гормоны, структурные. Белки, сократительные белки, антитела и т. д. Следует, однако, иметь в виду, что из-за несовершенства самой классификации, а также вследствие исключительного многообразия белков многие из отдельных белков не могут быть отнесены ни к одной из описываемых здесь групп.
Все белки принято делить на простые, или протеины (собственно белки), и сложные, или протеиды (комплексы белков с небелковыми соединениями). Простые белки являются полимерами только аминокислот; сложные, помимо остатков аминокислот, содержат также небелковые, так называемые простетические группы.
Среди простых белков (протеинов) различают альбумины (см.), глобулины (см.) и ряд других белков.
Альбумины - легко растворимые глобулярные белки (например, альбумины сыворотки крови или яичного белка); растворяются в воде и солевых растворах с выпадением в осадок лишь при насыщении раствора сульфатом аммония.
Глобулины отличаются от альбуминов нерастворимостью в воде и выпадением в осадок при полунасыщении раствора сульфатом аммония. Глобулины обладают более высоким, чем альбумины, молекулярным весом и иногда содержат в своем составе углеводные группировки.
К протеинам относятся и растительные белки - проламины (см.), встречающиеся обычно совместно с глютелинами (см.) в семенах злаков (рожь, пшеница, ячмень и др.), образуя основную массу клейковины. Эти белки растворимы в 70-80% спирте и нерастворимы в воде; они богаты остатками пролина и глутаминовой кислоты. К проламинам относятся также глиадин пшеницы, зеин кукурузы, гордеин ячменя.
Склеропротеины (протеинонды, альбуминоиды) - структурные белки, нерастворимые в воде, в разведенных щелочах, кислотах и солевых растворах. К ним относятся фибриллярные белки главным образом животного происхождения, весьма устойчивые к перевариванию пищеварительными ферментами. Эти белки подразделяют на белки соединительной ткани: коллаген (см.) и эластин (см.); белки покровов - волос, ногтей и копыт, эпидермиса- кератины (см.), для которых характерно высокое содержание серы в виде остатка аминокислоты - цистина; белки коконов и других секретов шелкоотделительных желез насекомых (например, паутины) - фиброин (см.), состоящие более чем наполовину из остатков глицина и аланина.
Особую группу протеинов составляют протамины (см.) - сравнительно низкомолекулярные белки основного характера (в отличие от альбуминов, глобулинов и других тканевых белков, имеющих изоэлектрическую точку обычно в слабокислой среде). Протамины встречаются в сперме некоторых рыб и других животных и состоят более чем наполовину из диаминомонокарбоновых кислот. Так, протамины сельди - клупеин и лосося - сальмин содержат около 80% аргинина. Другие протамины содержат, помимо аргинина, также лизин или лизин и гистидин.

Рис. 2. Общая схема биосинтеза белка. Аминокислоты (1), взаимодействуя с АТФ, активируются, образуя аминоациладенилаты (2); последние под действием фермента аминоацил-тРНК-синтетазы соединяются с транспортными РНК, или тРНК (3), и в виде комплекса аминоацил-тРНК (4) поступают в рибосомы, соединенные с мРНК, или полисомы (5). Полисомы образуются путем присоединения к мРНК сначала малой субчастицы (6), а затем и большой субчастицы (7) рибосом. В рибосоме (8), соединенной с мРНК, к мРНК присоединяются две аминоацил-тРНК, в результате чего между ними образуется пептидная связь. Таким образом происходит рост полипептидной цепи (9), которая освобождается по завершении ее синтеза (10) и далее трансформируется в белок (11).
Биосинтез белков протекает во всех клетках живых организмов и обеспечивает обновленце белков организма, процессы обмена веществ и их регуляцию, а также рост и дифференцировку органов и тканей. Белки синтезируются в тканях из свободных аминокислот при участии нуклеиновых кислот (см.). Процесс биосинтеза белков протекает с потреблением энергии, аккумулированной в виде АТФ (см. Аденозинфосфорные кислоты). При биосинтезе белков обеспечивается образование определенных белков строго специфической структуры, которая закодирована в структурных генах (цистронах) дезоксирибонуклеиновой кислоты, находящейся главным образом в хроматине ядер клеток (см. Генетический код). Информация, определяющая первичную структуру белков, передается на особый тип рибонуклеиновых кислот (РНК), называемых информационными, или матричными, РНК (мРНК), в виде комплементарной последовательности нуклеотидов. Этот процесс получил название транскрипции. мРНК соединяется с рибосомами (см.), представляющими собой рибонуклеопротеидные гранулы, более чем наполовину состоящие из особой рибосомной РНК (рРНК), синтезируемой также на специальных цистронах (генах) ДНК. Рибосомы состоят из двух субчастиц, на которые они способны обратимо диссоциировать при понижении концентрации ионов магния. Большая и малая субчастицы рибосом содержат но одной молекуле РНК с молекулярной массой соответственно около 1,7×10 6 и 0,7×10 6 и по нескольку десятков молекул белков. Соединившись с рибосомами, мРНК образует полирибосомы, или полисомы, на которых и происходит синтез полипептидных цепочек, образующих первичную структуру белков. Прежде чем соединиться с рибосомами аминокислоты активируются, затем соединяются с низкополимерными РНК-переносчиками, или транспортными РНК (тРНК) в виде комплексов, с которыми они и поступают в рибосомы. Общая схема биосинтеза белков представлена на рис. 2.
Активация аминокислот происходит при взаимодействии их с АТФ с образованием аминоациладенилата и освобождением пирофосфата: аминокислота + АТФ = аминоациладенилат + пирофосфат. Аминоациладенилат представляет собой смешанный ангидрид, образованный остатком фосфорной к-ты аденозинмонофосфата и карбоксильной группой аминокислоты, и является активированной формой аминокислоты. С аминоациладенилата остаток аминокислоты переносится на тРНК, специфичную для каждой аминокислоты, и в виде аминоацил-тРНК поступает в рибосомы. Образование аминоациладенилата и перенос аминокислотного остатка на тРНК катализируются одним и тем же ферментом (аминоациладенилатсинтетазой, или аминоацил-тРНК-синтетазой), строго специфичным для каждой аминокислоты и каждой тРНК. Все тРНК имеют сравнительно небольшой молекулярный вес (около 25 000) и содержат около 80 нуклеотидов. Они имеют крестообразную конфигурацию, напоминающую клеверный лист, причем нуклеотидная цепь образует двунитчатую структуру, удерживаемую комплементарными основаниями, и переходит в однонитчатую только в области петель. Начало нуклеотидной цепи, обычно представленное 5"-гуаниловым нуклеотидом, находится вблизи концевой, часто обменивающейся группировки из двух остатков цитидиловой кислоты и аденозина со свободной 3"-OH-группой, к которой и присоединяется остаток аминокислоты. На петле, находящейся у противоположного конца молекулы тРНК, имеется триплет оснований, комплементарный к триплету, кодирующему данную аминокислоту (кодону), и называемый антикодоном. Нуклеотидная последовательность многих тРНК уже установлена, известна и их полная структура.
Определенная последовательность аминокислот в первичной структуре синтезируемой полипептидной цепи обеспечивается информацией, записанной в последовательности нуклеотидов мРНК, отражающей соответствующую последовательность в цистронах ДНК. Каждая аминокислота кодируется определенными триплетами нуклеотидов мРНК. Эти триплеты (кодоны) представлены в табл. 2. Их расшифровка позволила установить нуклеотидный код РНК, или аминокислотный код, то есть способ, при помощи которого происходит трансляция, или перевод информации, записанной в последовательности нуклеотидов РНК в первичную структуру белков, или последовательность аминокислотных остатков в полипептидной цепи.
Таблица 2. РНК-АМИНОКИСЛОТНЫЙ КОД
|
Первый нуклеотид кодона (с 5"-конца) |
Второй нуклеотид кодона |
Третий нуклеотид кодона (с 3’-конца) |
|||
|
Фен |
Сер |
Тир |
Цис |
||
|
Фен |
Сер |
Тир |
Цис |
||
|
Лей |
Сер |
УАА |
УГА |
||
|
Лей |
Сер |
УАГ |
Три |
||
|
Лей |
Про |
Гис |
Арг |
||
|
Лей |
Про |
Гис |
Арг |
||
|
Лей |
Про |
Глн |
Арг |
||
|
Лей |
Про |
Глн |
Арг |
||
|
Иле |
Тре |
Асн |
Сер |
||
|
Иле |
Тре |
Асн |
Сер |
||
|
Иле |
Тре |
Лиз |
Арг |
||
|
Мет |
Тре |
Лиз |
Арг |
||
|
Вал |
Ала |
Асп |
Гли |
||
|
Вал |
Ала |
Асц |
Гли |
||
|
Вал |
Ала |
Глу |
Гли |
||
|
Вал |
Ала |
Глу |
Гли |
||
Примечание: У - уридиловая кислота, Ц - цитидиловая кислота, А - адениловая кислота, Г - гуаниловая кислота. Три буквы обозначают соответствующий аминокислотный остаток: напр.. Фен - фенилаланин. Иле - изолейцин, Глу - глутаминовая кислота, Глн - глутамин и т. п. Триплеты УАА, УАГ, УГА не кодируют аминокислот, но определяют терминацию полипептидной цепи.
Как видно из таблицы, из 64 возможных триплетов (61 кодируют определенные аминокислоты, то есть являются «смысловыми». Три триплета - УДА, УАГ и УГА - не кодируют аминокислот, однако их роль заключается в завершении (терминации) синтеза растущей полипептидной цепочки. Код является вырожденным, то есть почти все аминокислоты кодируются более чем одним триплетом нуклеотидов. Так, 3 аминокислоты - лейцин, аргинин и серии - кодируются шестью кодонами, 2 - метионин и триптофан - имеют только по одному кодону, а остальные 15 - от 2 до 4. Процесс трансляции осуществляется при помощи тРНК, нагруженных аминокислотами. Аминоацил-тРНК присоединяется своим комплементарным триплетом (антикодоном) к кодону мРНК в рибосоме. К соседнему кодону мРНК присоединяется другая аминоацил-тРНК. Первая тРНК при этом присоединяет свой аминокислотный остаток карбоксильным концом к аминогруппе второй аминокислоты, с образованием дипептида, а сама освобождается и отделяется от рибосомы. Далее, по мере продвижения рибосомы но цепи мРНК от 5"-конца к З"-концу, присоединяется третья аминоацил-РНК; происходит соединение дипептида карбоксильным концом с аминогруппой третьей аминокислоты с образованием трипептида и освобождением второй тРНК и так до тех пор, пока рибосома не пройдет весь участок, кодирующий данный белок на мРНК, соответствующий цистрону ДНК. Затем происходит терминация синтеза белков, и образовавшийся полипептид освобождается от рибосомы. За первой рибосомой в полисоме следует вторая, третья и т. д., которые последовательно считывают информацию на одной и той же нити мРНК в полисоме. Таким образом, рост полипептидной цепи происходит с N-конца к карбоксильному (С-) концу. Если подавить синтез белков, например, при помощи антибиотика пуромицина, то можно получить недостроенные полипептидные цепи с незавершенным на разных этапах С-концом. Аминоацил-тРНК присоединяется сначала к малой рибосомной субчастице, а затем переносится на большую субчастицу, на которой и происходит рост полипептидной цепочки. Согласно гипотезе А. С. Спирина во время работы рибосомы при биосинтезе белков происходит повторяющееся смыкание и размыкание субчастиц рибосом. Для воспроизведения синтеза белков вне организма, помимо рибосом, мРНК и аминоацил-тРНК, необходимо присутствие гуанозинтрифосфата (ГТФ), который расщепляется до ГДФ и снова регенерирует в процессе роста поли пептидной цепи. Необходимо также присутствие нескольких белковых факторов, выполняющих, по-видимому, ферментативную роль. Эти так называемые трансферные факторы взаимодействуют друг с другом и для своей активности требуют присутствия сульфгидрильных групп и ионов магния. Помимо собственно трансляции (то есть роста полипептидной цепи в определенной последовательности, соответствующей структурному гену ДНК и передаваемой последовательностью нуклеотидов в мРНК), особую роль играет начало (или инициация) трансляции и завершение (или терминация) ее. Инициация белкового синтеза в рибосоме, по крайней мере в бактериях, начинается с особых кодонов - инициаторов в мРНК - АУГ и ГУГ. Сначала с таким кодоном связывается малая субчастица рибосомы затем к ней присоединяется формилметионил-тРНК, с которой и начинается синтез полипептидной цепи. В силу особых свойств этой аминоацил-тРНК она способна переноситься на большую субчастнцуг подобно пептидил-тРНК, и таким образом начинать рост полипептидной цепи. Для инициации необходимы ГТФ и белковые факторы инициации (известно три). Терминация роста полипептидной цепи происходит на «бессмысленных» кодонах УАА, УАГ или УГА. По-видимому, эти кодоны связываются с особым белковым фактором терминации, который в присутствии еще одного фактора способствует освобождению полипептида.
Компоненты системы биосинтеза белков синтезируются главным образом в клеточном ядре. На матрице ДНК в процессе транскрипции происходит синтез всех типов РНК. участвующих: в этом процессе: рРНК, мРНК и тРНК. Так, рРНК и мРНК синтезируются в виде очень больших молекул и еще в клеточном ядре проходят процесс «созревания», в ходе которого часть (весьма значительная для мРНК) молекул отщепляется и подвергается распаду, не выходя в цитоплазму, а функционирующие молекулы, являющиеся частью первоначально синтезированных, поступают в цитоплазму к местам белкового синтеза. Прежде чем попасть в состав полисом, мРНК, по-видимому, с момента синтеза связывается с особыми белковыми частицами, «информоферами», и в виде рибонуклеопротеидного комплекса переносится к рибосомам. Рибосомы, очевидно, также «дозревают» в цитоплазме, часть белков присоединяется к предшественникам рибосом, выходящим из ядра, уже в цитоплазме. Следует отметить, что у низших, безъядерных организмов (прокариотов), к которым относятся бактерии, сине-зеленые водоросли и вирусы, имеются некоторые отличия от высших организмов в компонентах системы биосинтеза белков, и в особенности в его регуляции. Рибосомы у прокариотов несколько меньше по размерам и отличаются по составу, процесс транскрипции и трансляции непосредственно связан в одно целое. Вместе с тем у высших ядерных организмов (эукариотов) образование РНК происходит и в органеллах цитоплазмы, митохондриях и хлоропластах (у растений), обладающих собственной системой синтеза белка и собственной генетической информацией в виде ДНК. По своему устройству система белкового синтеза в митохондриях и хлоропластах аналогична таковой у прокариотов и существенно отличается от системы, имеющейся в ядре и цитоплазме высших животных и растений.
Регуляция биосинтеза белков представляет весьма сложную систему и позволяет клетке быстро и четко реагировать на изменения в окружающей клетку среде путем прекращения или индукции синтеза различных белков, часто обладающих ферментативной активностью. У бактерий подавление синтеза белков осуществляется главным образом при помощи особых белков - репрессоров (см. Оперон), синтезируемых специальными генами-регуляторами. Взаимодействие репрессора с метаболитом, поступающим из среды или синтезируемым в клетке, может подавить или, наоборот, активировать его, регулируя таким образом синтез одного белка или нескольких взаимосвязанных белков, в особенности ферментов, синтезирующихся также взаимосвязанно на одном опероне. У высших организмов в процессе дифференцировки ткани теряют способность к синтезу ряда белков и специализируются на синтезе меньшего числа белков, необходимых для функции данной ткани, например мышц. Такое блокирование синтеза ряда белков происходит, по-видимому, на уровне генома (см.) при помощи ядерных белков - гистонов (см.), связывающих нефункциональные участки ДНК. Однако при регенерации, злокачественном росте и других процессах, связанных с дедифференцировкой, такие заблокированные участки могут дерепрессироваться и поставлять мРНК для синтеза необычных для данной ткани белков. Тем не менее и у высших организмов имеет место регуляция синтеза белков в ответ на те или иные стимулы. Так, действие ряда гормонов заключается в индукции синтеза белков в ткани, являющейся «мишенью» данного гормона. Такая индукция, по-видимому, происходит путем связывания гормона с особым белком данной ткани и активацией гена посредством образованного комплекса.
Процесс биосинтеза белков и его регуляция требуют чрезвычайной четкости, точности и слаженности работы всех компонентов системы. Даже небольшие нарушения этой точности приводят к нарушению первичной структуры белков и тяжелым патологическим последствиям. Генетические нарушения, например, замена или потеря одного нуклеотида в структурном гене, приводят к синтезу измененного белка, нередко лишенного биологической активности. Такие изменения лежат в основе врожденных нарушений обмена веществ, к которым, по существу, относятся все наследственные болезни (см.). С другой стороны, целый ряд белков и ферментов может различаться не только у разных биологических видов, но и у разных индивидуумов, сохраняя при этом свою биологическую активность. Нередко такие белки обладают разными иммунологическими и электрофоретическими свойствами. В популяциях человека описаны многие примеры так называемого полиморфизма белков, когда у разных индивидуумов, а иногда и у одного и того же индивидуума можно обнаружить два или несколько неодинаковых белков, обладающих одной и той же функцией, как, например, гемоглобин (см.), гаптоглобин (см.) и некоторые другие.
Белки в питании
Среди многочисленных пищевых веществ белкам принадлежит наиболее важная роль. Они являются источниками незаменимых аминокислот и так называемого неспецифического азота, необходимых для синтеза белков человеческого организма. Выраженная недостаточность белков в питании приводит к тяжелым нарушениям функции организма (см. Алиментарная дистрофия). От уровня снабжения белками в большой степени зависит состояние здоровья, физического развития и- работоспособности человека, а у детей раннего возраста в определенной мере и умственное развитие. Если учесть все производимые для питания растительные и животные белки, то в среднем на каждого жителя Земли придется около 58 г в день. В действительности более половины населения, особенно развивающихся стран, не получает этого количества белка. Глобальный дефицит пищевого белка должен быть отнесен к числу наиболее острых экономических и социальных проблем современности (см. Кризис белковый). В связи с этим установление оптимальных уровней содержания белка в пищевых рационах приобретает первостепенную важность.
В наибольших количествах белки требуются в периоды интенсивного роста. Однако и в организме, достигшем зрелости, процессы жизнедеятельности связаны с непрерывной тратой белковых веществ и, следовательно, необходимостью воспол нения этих потерь с пищей. В соответствии с рекомендациями Экспертной группы ФАО/ВОЗ расчет потребности в белковом азоте следует проводить по формуле: R=1,1(U b +F b +S+G), где R - потребность в белковом азоте; U b - выделение азота с мочой; F b - выделение азота с калом; S - потеря азота за счет десквамации эпидермиса, роста волос, ногтей, выделения азота с потом при неинтенсивном потении; G - удержание азота в процессе роста (расчет ведется на 1 кг массы в день).
Коэффициент 1,1 отражает добавочные траты белков (в среднем 10%), возникающие в результате стрессовых реакций и неблагоприятных воздействий на организм. Границы индивидуальных вариаций потребностей в белках принимаются равными ±20%. Официальные рекомендации экспертной группы ФАО/ВОЗ отражены в табл. 3.
Таблица 3. СРЕДНЕСУТОЧНАЯ ПОТРЕБНОСТЬ В БЕЛКАХ (при условии его полного усвоения)*
|
Возраст (в годах) |
Потребность (в г на 1 кг массы тела в день) |
||
|
средняя |
-20% |
+20% |
|
|
Дети |
|||
|
1-3 |
0,88 |
0,70 |
1,06 |
|
4-6 |
0,81 |
0,65 |
0,97 |
|
7-9 |
0,77 |
0,62 |
0,92 |
|
10-12 |
0,72 |
0,58 |
0,86 |
|
Подростки |
|||
|
13-15 |
0,70 |
0,56 |
0,84 |
|
16-19 |
0,64 |
0,51 |
0,77 |
|
Взрослые |
0,59 |
0,47 |
0,71 |
- Величина потребности в азоте умножена на коэффициент 6,25.
Очевидно, что приведенные величины но соответствуют оптимальному снабжению человека белками и должны быть отнесены к минимальному уровню их содержания в рационе, при несоблюдении которого неизбежно сравнительно быстрое развитие серьезных последствий белковой недостаточности. Фактическое потребление белков в большинстве экономически развитых стран в 1,5 и даже 2 раза выше приведенных цифр. Согласно концепции сбалансированного питания, оптимальная потребность человека в белках зависит от многих факторов, включая физиологические особенности организма, качественную характеристику пищевых белков и содержание в рационе других пищевых веществ.
В СССР величины потребностей населения в белках зафиксированы в официально утверждаемых Министерством здравоохранения физиологических нормах питания, которые периодически пересматриваются и уточняются. Физиологические нормы питании являются средними ориентировочными величинами, отражающими оптимальные потребности отдельных групп населения в основных пищевых веществах и энергии (табл. 4).
|
Детское население |
||
|
возраст |
потребление белков |
|
|
всего |
животных |
|
|
0 - 3 мес . |
||
|
4- 6 мес . |
||
|
6- 12 мес . |
||
|
1 - 1,5 года |
||
|
1,5- 2 года |
||
|
3- 4 года |
||
|
5- 6 лет |
||
|
7-10 лет |
||
|
11 - 13 лет |
||
|
14- 17 лет (юноши) |
||
|
14- 17 лет (девушки) |
||
|
Взрослое население |
|||||
|
группы по характеру труда |
(в годах |
мужчины |
женщины |
||
|
потребление белков |
потребление белков |
||||
|
всего |
живот ных |
всего |
живот ных |
||
|
Труд , не связанный с физическими напряжениями |
18- 40 |
||||
|
Механизированный труд и сфера обслуживания с невысокой физической нагрузкой |
|||||
|
40 - 60 |
|||||
|
Механизированный труд и сфера обслуживания со значительной нагрузкой |
18 - 40 |
||||
|
Механизированный труд с большой физ . нагрузкой |
|||||
|
Пенсионный возраст |
60- 70 |
||||
|
Свыше |
|||||
|
Студенты |
|||||
|
Беременные 5 -9 мес . |
|||||
|
Кормящие |
|||||
В них предусматриваются дифференциация потребностей в белках, в зависимости от пола, возраста, характера труда и т. д. Рекомендуемые величины рассчитаны на основании изучения особенностей белкового обмена и азотистого баланса у соответствующих групп населения, причем они значительно выше минимальных потребностей в белках, необходимых для поддержания азотистого равновесия. Избыток белков необходим дли обеспечения дополнительных трат организма, связанных с физическими и нервными нагрузками, неблагоприятными воздействиями внешней среды, а также для поддержания оптимального иммунологического статуса. Специально выделены в нормах величины потребления наиболее ценных белков животного происхождения.
Физиологические нормы питания являются основой планирования производства тех или иных пищевых продуктов. При оценке полезности отдельных белковых продуктов учитывается их аминокислотный состав, степень перевариваемости ферментами пищеварительного тракта и интегральные показатели усвояемости, установленные в результате биологических экспериментов. На практике с определенной степенью условности белковые продукты делят на две группы. К первой относят продукты животного происхождения: молоко, мясо, яйца, рыбу, белки которых легко и полностью усваиваются организмом человека; ко второй - большинство продуктов растительного происхождения, в частности пшеницу, рис, кукурузу и другие злаковые, белки которых усваиваются организмом не полностью. Условность подобного деления подчеркивается высокой биологической ценностью ряда белков растительного происхождения (картофеля, гречихи, сои, подсолнечника) и низкой биологической ценностью белков некоторых животных продуктов (желатины, кожи, сухожилий и др.). Причины низкой усвояемости фибриллярных белков (кератина, эластина и коллагенов) заключаются в особенностях их третичной структуры и трудности переваривания ферментами пищеварительного тракта. С другой стороны, усвоение ряда белков растительного происхождения может зависеть от структуры растительных клеток и возникающих трудностей в контактировании белков с пищеварительными ферментами.
Полнота использования отдельных белков человеком или их биологическая ценность и первую очередь определяются степенью соответствия их аминокислотного состава дифферинцированным потребностям организма и в какой-то степени аминокислотному составу тела. Огромное разнообразие встречающихся в природе белков в основном построено из 20 аминокислот, 8 из них (триптофан, лейцин, изолейцин, валин, треонин, лизин, метионин и фенилаланин) незаменимы для человека, так как не могут быть синтезированы в тканях организма (см. Аминокислоты). Дли маленьких детей девятой незаменимой аминокислотой является гистидин. Остальные аминокислоты относятся к числу заменимых и могут расцениваться в питании главным образом как поставщики неспецифического азота. Установлено, что лучшее усвоение белков пищи достигается при сбалансировании ее аминокислотного состава с «идеальными» аминокислотными шкалами. В качестве подобной шкалы в 1957 году была предложена так называемая предварительная аминокислотная шкала ФАО. Позднее было доказано, что содержание в ней ряда аминокислот, особенно триптофана и метионина, определено не вполне точно. В соответствии с результатами биологических исследований в качестве оптимальных в последние годы рекомендованы шкалы аминокислотного состава белков куриных яиц и женского молока. Белки этих двух продуктов самой природой предназначены для питания развивающихся организмов и практически полностью утилизируются как в опытах на экспериментальных животных, так и при использовании в питании детей раннего возраста.
Для определения соответствия аминокислотного состава белков потребностям человека предложен ряд индексов, каждый из которых имеет лишь ограниченную ценность. В их числе следует упомянуть индекс Н/О, отражающий отношение суммы незаменимых аминокислот (Н в мг) к общему содержанию азота белков (О в г), который помогает определению соотношении азота незаменимых, или эссенциальных, аминокислот и неспецифического азота. Чем ниже величина Н/О тем выше содержание неспецифического азота. Для белков молока и яиц этот индекс сравнительно высок - 3,1-3,25, для мяса - 2,79-2,94; для пшеницы - 2. Большое значение придается показателю аминокислотного скора, позволяющему получить более полное суждение о биологической ценности белка на основании его хим. состава.
Метод скора основан на подсчете в исследуемом продукте процента обеспечения каждой из незаменимых аминокислот по сравнению с идеальными аминокислотными шкалами.
С этой целью для каждой из эссенциальных аминокислот исследуемого белка рассчитывается величина I иссл, равная А иссл /Н иссл, отражающая отношение каждой незаменимой аминокислоты (А в мг) к сумме незаменимых аминокислот (Н в г); полученная цифра сопоставляется с величиной I ст, равной А ст /Н ст для той же аминокислоты, рассчитанной по стандартной шкале. В результате деления величин Iиссл на Iст и умножения на 100 получают показатель аминокислотного скора для каждой из незаменимых аминокислот. Лимитирующей биологическую ценность исследуемого белка является аминокислота, показатель аминокислотного скора для которой является наименьшим. В качестве стандартных шкал наряду с предварительной шкалой ФАО используют аминокислотные шкалы куриных яиц и женского молока (табл. 5).
Таблица 5. СТАНДАРТНЫЕ АМИНОКИСЛОТНЫЕ ШКАЛЫ
|
Аминокислоты |
Отношение незаменимой аминокислоты в мг к 1 г суммы незаменимых аминокислот (А /Н ) |
|||||
|
женское молоко |
куриные яйца |
женское молоко |
куриные яйца |
|||
|
Изолейцин |
||||||
|
Лейцин |
||||||
|
Лизин |
||||||
|
Сумма ароматических аминокислот : |
||||||
|
фенилаланин |
||||||
|
тирозин |
||||||
|
Сумма серосодержащих аминокислот : |
||||||
|
цистин |
||||||
|
метионин |
||||||
|
Треонин |
||||||
|
Триптофан |
||||||
|
Валин |
||||||
|
Сумма незаменимых аминокислот |
||||||
В соответствии с показателями аминокислотного скора (табл. 6) наименьшей биологической ценностью обладают белки ряда злаковых, особенно пшеницы (50%; лимитирующие аминокислоты - лизин и треонин); кукурузы (45%; лимитирующие аминокислоты - лизин и триптофан); проса (60%; лимитирующие аминокислоты - лизин и треонин); гороха (60%; лимитирующие аминокислоты - метионин и цистин). Показатель аминокислотного скора лимитирующей аминокислоты устанавливает предел использования азота данного вида белка для пластических целей. Избыток других содержащихся в белке аминокислот может использоваться только как источник неспецифического азота или для энергетических нужд организма. Метод изучения аминокислотного состава является одним из основных способов оценки качества белков. Обычно он позволяет получить показатели усвояемости, близкие по отношению к результатам более длительных и дорогостоящих методов биологического определения ценности белков. В то же время установление в ряде случаев достоверных расхождений между указанными показателями заставляет прибегать при исследовании новых белковых продуктов к интегральным методам биол. оценки как на лабораторных животных, так и непосредственно на людях. Эти методы основаны на изучении в балансовых опытах полноты использования отдельных белков растущими животными (показатель белковой эффективности рациона), соотношения удерживаемого организмом азота к азоту, адсорбированному из кишечника (показатель биол. ценности), отношения адсорбированного азота к общему азоту пищи (показатель истинной переваримости) и т. п. При постановке исследований по изучению биол, ценности белка обязательным является достаточно калорийное обеспечение рациона, его сбалансирование по всем незаменимым факторам питания(см. Сбалансированное питание) и сравнительно низкий уровень белков - в пределах 8- 10% от общей калорийности (см. Обмен веществ и энергии). Сопоставление показателей аминокислотного скора и утилизации белка, определенной в опытах на экспериментальных животных для некоторых продуктов, представлено в табл. 6.
Таблица 6. СОПОСТАВЛЕНИЕ ПОКАЗАТЕЛЕЙ АМИНОКИСЛОТНОГО СКОРА И УТИЛИЗАЦИИ БЕЛКА
|
Продукты |
Аминокислотный скор |
Лимитирующие аминокислоты |
Показа -тели утилизации белка |
||
|
по шкале ФАО |
по женскому молоку |
по куриным яйцам |
|||
|
Молоко коровье |
|||||
|
Яйца |
|||||
|
Казеин |
|||||
|
Яичный альбумин |
Триптофан |
||||
|
Мясо говяжье |
|||||
|
Сердце говяжье |
|||||
|
Печень говяжья |
|||||
|
Почки говяжьи |
|||||
|
Свинина (вырезка ) |
|||||
|
Рыба |
Триптофан |
||||
|
Овес |
Лизин |
||||
|
Рожь |
Треонин |
||||
|
Рис |
Лизин |
||||
|
Кукурузная мука |
Триптофан |
||||
|
Просо |
во |
Лизин |
|||
|
Сорго |
|||||
|
Пшеничная мука |
|||||
|
Пшеничные зародыши |
|||||
|
Пшеничный глютент |
Лизин |
||||
|
Арахисовая мука |
|||||
|
Соевая мука |
|||||
|
Семена кунжута |
Лизин |
||||
|
Семена подсолнечника |
|||||
|
Семена хлопчатника |
|||||
|
Картофель |
|||||
|
Горох |
|||||
|
Батат (сладкий картофель ) |
|||||
|
Шпинат |
|||||
|
Кассава |
|||||
Важным преимуществом биологических методов оценки белков является их интегральность, дающая возможность учитывать весь комплекс свойств продуктов, влияющих на усвояемость входящих в них белков. Изучая биологическую ценность отдельных белков, не следует забывать, что практически во всех рационах питания используются не отдельные белки, а их комплексы, причем, как правило, различные белки взаимно дополняют друг друга, обеспечивая некоторые средние показатели усвоения белкового азота. При достаточно разнообразных смешанных диетах показатель переваримости белков рационов питания сравнительно постоянен и приближается к 85%, что нередко используется при проведении практических расчетов.
Рис. 2. Реакция Даниэлли на белки, содержащие тирозин, триптофан, гистидин в ушке сердца.

В основе гистохимических методов выявления белков лежат, как правило, биохимические методы, приспособленные для определения белков в тонких тканевых срезах. Следует иметь в виду, что биохимическая реакция может быть использована как гистохимическая в том случае, если продукт реакции имеет устойчивую цветную окраску, выпадает в осадок и не обладает выраженной склонностью к диффузии. Гистохимические методы выявления белков в тканях базируются на выявлении определенных аминокислот, входящих в состав белков (например, реакция Миллона на тирозин, реакция Сакагуши на аргинин, реакция Адамса на триптофан, реакция тетразониевого сочетания на гистидин, тирозин, триптофан и т. д.), на выявлении определенных химических групп (NH 2 =,COOH - ,SH=,SS = и т. п.), на применении некоторых физико-химических методов (цветн. рис. 1-3), определении изоэлектрической точки и т. д. Наконец, выяснить наличие в тканевом срезе некоторых аминокислот можно косвенным путем, определив наличие в тканях ферментов, связанных с этими аминокислотами (например, оксидазы D-аминокислот). Некоторые простые белки (коллаген, эластин, ретикулин, фибрин) выявляются в срезах с помощью многочисленных гистологических методов, среди которых предпочтительными являются так называемые полихромные методы (метод Маллори и его модификации, орсеинпикрофуксиновый метод Ромейса и др. Выявляются белки и при использовании методов люминесцентной микроскопии. Локализацию белков в тканях (миозинов, альбуминов, глобулинов, фибрина и т. д.) можно получить при помощи метода меченых антител по Кунсу и др. Эти методы и их модификации позволяют достаточно точно идентифицировать и определить локализацию отдельных белков, отличающихся друг от друга содержанием тех или иных аминокислот. Разрабатываются методы количественного определения белков, например, метод определения белков непрямой реакцией меченых антител, а также определения SH-групп по методу Барнетта и Зелигмана (см. Аминокислоты , гистохимические методы выявления аминокислот). Все упомянутые выше методы выявления белков в тканях обладают достаточной специфичностью и дают вполне достоверные результаты. Фиксация тканевого материала при использовании названных методов различна. Наиболее подходящими фиксаторами в большинстве случаев следует считать этиловый или метиловый спирт, обезвоженный ацетон, смесь этилового спирта с формалином, раствор трихлоруксусной кислоты на спирте, в некоторых случаях (для протеидов передней доли гипофиза) применяется формалин. Выбор фиксатора зависит от метода, время фиксации - от общего количества и характера ткани. Можно использовать криостатные или парафиновые срезы.
Радиоактивные белки
Радиоактивные белки - белковые вещества, в молекуле которых содержится один или несколько атомов радиоактивных изотопов каких-либо элементов. При радиоактивном мечении белков необходимо обеспечить прочность и возможно большую сохранность белковой молекулы. В качестве радиоактивной метки белков для биохимических экспериментальных исследований используются главным образом изотопы 3 Н и 14 С; при получении радиофармацевтических препаратов на основе белков применяют изотопы йода - 125 I и 131 I, а также изотопы 111 In, 113m In , 99м Tc и др. Введение изотопов йода в белки основано на электрофильном замещении водорода на йод в фенольном ядре тирозина молекулы белка или пептида. Меченый белок очищают от несвязанного йодида и других примесей (путем гельфильтрации, диализа, адсорбции, ионообмена, изоэлектрического осаждения и др.). Если белки не содержит тирозина, для проведения йодирования в него вводят содержащие радиоактивный йод заместители, или используют тирозинсодержащие аналоги, или же прибегают к метке другими радиоактивными изотопами (см.).
Радиоактивные белки имеют важное значение в изучении катаболизма и метаболизма белковых веществ в экспериментальных биохимических исследованиях. Кроме того, их используют в радиоизотопной диагностике in vivo и in vitro при изучении функционального состояния многих органов и систем организма в случае различных заболеваний. В исследованиях in vivo наибольшее применение находит альбумин сыворотки крови человека, меченный радиоактивными изотопами йода (125 I и 131 I), а также получаемые на его основе путем тепловой денатурации и агрегации с той же меткой микро-и макроагрегаты альбумина. С помощью меченого альбумина могут быть определены показатели гемодинамики и регионального кровообращения, объем циркулирующей крови и плазмы, произведено сканирование сердца и крупных сосудов (см. Сканирование), а также опухолей головного мозга. Микроагрегаты альбумина используют для сканирования печени и желудка, определения кровотока печени, а макроагрегаты - для сканирования легких.
Радиоактивные белки нашли широкое применение при определении микроколичеств гормонов, ферментов и других белковых веществ в тканях и средах организма животных и человека в исследованиях in vitro.
Библиография: Белки, под ред. Г. Нейрата и К. Бэйли, пер. с англ., т. 1-3, М., 1956 -1959, библиогр.; Биосинтез белка и нуклеиновых кислот, под ред. А. С. Спирина, М., 1965; Гауровнц Ф. Химия и функции белков, пер. с англ.. М., 1965; Ичас М. Биологический код, пер. с англ., М., 1971; Киселев Л. Л. и др. Молекулярные основы биосинтеза белков. М., 1971; Поглааов Б. Ф. Структура и функции сократительных белков, М., 1965; Спирин А. С. и Гаврилова Л. П. Рибосома, М., 1971; Химия и биохимия нуклеиновых кислот, под ред. И. Б. Збарского и С. С. Дебова, Л., 1968; Advances in protein chemistry, ed. by M. L. Anson a. J. T. Edsall, v. 1-28, N. Y., 1944-1974; Hess G. P. a. R up ley J. A. Structure and function of proteins, Ann. Rev. Biochcm., v. 40, p. 1013, 1971; In vitro procedures with radioisotopes in mcdlcinc, Proceedings of the symposium, Vienna, 1970; M a r g-l(n A. a. Nerrif ieldR.B. Chemical synthesis of peptides and proteins, Ann. Rev. Biochem.,v. 39,p. 841, 1970; Proteins, composition, structure, and function, ed. by H. Neurath, v. 1 - 5, N. Y.-L., 1963- 1970.
Б. в питании - Лавров Б. A. Учебник физиологии питания, с. 92, М., 1935; Молчанова О. П. Значение белка в питании для растущего и взрослого организма, в кн.: Вопр. пит., под ред. О. П. Молчановой, в. 2, с. 5, М., 1950; П о к р овский А. А. К вопросу о потребностях различных групп населения в энергии и основных пищевых веществах, Вестн. АМН СССР, № 10, с. 3, 1966, библиогр.; он же, Фиэиолого-биохимические основы разработки продуктов детского питания, М., 1972; Energy
Гистохимические методы выявления Б. в тканях - Кисели Д. Практическая микротехника и гистохимия, пер. с веягер., с. 119, 152, Будапешт» 1962; Л и л-л я р. Патогистологическая техника и Фактическая гистохимия, пер. с англ., с. 509, М., 1969; П и р с Э. Гистохимия, пер. е англ.. М., 1962; Принципы и методы гп-гго-цитохимического анализа в патологии, аод ред. А. П. Авцына и др., с. 238, JI., ".971; Р е а г s е A. G. Е. Histochemistry, т. 1-2, Edinburgh - L., 1969-1972.
И. Б. Збарский; А. А. Покровский (пит.), В. В. Седов (рад.), Р. А. Симакова (гист.).
Более 4 млрд лет назад на Земле из маленьких неорганических молекул непостижимым образом возникли белки, ставшие строительными бло-ками живых организмов. Своим бес-конечным разнообразием всё живое обязано именно уникальным молеку-лам белка, и иные формы жизни во Вселенной науке пока неизвестны.
Белки, или протеины (от греч. «протос» — «первый»), — это природ-ные органические соединения, кото-рые обеспечивают все жизненные процессы любого организма. Из бел-ков построены хрусталик глаза и па-утина, панцирь черепахи и ядовитые вещества грибов... С помощью белков мы перевариваем пищу и боремся с болезнями. Благодаря особым белкам по ночам светятся светлячки, а в глу-бинах океана мерцают таинствен-ным светом медузы.
Белковых молекул в живой клетке во много раз больше, чем всех других (кроме воды, разумеется!). Учёные вы-яснили, что у большинства организ-мов белки составляют более полови-ны их сухой массы. И разнообразие видов белков очень велико — в одной клетке такого маленького организма, как бактерия Escherichia сой" (см. до-полнительный очерк «Объект иссле-дования — прокариоты»), насчиты-вается около 3 тыс. различных белков.
Впервые белок был выделен (в ви-де клейковины) в 1728 г. итальянцем Якопо Бартоломео Беккари (1682— 1766) из пшеничной муки. Это собы-тие принято считать рождением хи-мии белка. С тех пор почти за три столетия из природных источников получены тысячи различных белков и исследованы их свойства.
БИОЛОГИЧЕСКИЕ «БУСЫ»
Молекула белка очень длинная. Хими-ки называют такие молекулы поли-мерными (от греч. «поли» — «много» и «мерос» — «часть», «доля»). Действи-тельно, длинная молекула полимера состоит из множества маленьких мо-лекул, связанных друг с другом. Так нанизываются на нить бусинки в ожерелье. В полимерах роль нити иг-рают химические связи между бусин-ками-молекулами.
Секрет белков спрятан в особен-ностях этих самых бусинок. Боль-шинство полимеров не принимает устойчивой формы в пространстве, уподобляясь тем же бусам, у которых и не может быть пространственной структуры: повесишь их на шею — они примут форму кольца или овала, положишь в коробку — свернутся в клубок неопределённой формы. А те-перь представим себе, что некоторые бусинки могут «слипаться» друг с другом. Например, красные притяги-ваются к жёлтым. Тогда вся цепочка примет определённую форму, обязан-ную своим существованием «слипа-нию» жёлтых и красных бусинок
Нечто подобное происходит и в белках. Отдельные маленькие моле-кулы, входящие в состав белка, обла-дают способностью «слипаться», так как между ними действуют силы при-тяжения. В результате у любой белко-вой цепи есть характерная только для неё пространственная структура. Именно она определяет чудесные свойства белков. Без такой структуры они не могли бы выполнять те функ-ции, которые осуществляют в живой клетке.
При длительном кипячении бел-ков в присутствии сильных кислот или щелочей белковые цепи распада-ются на составляющие их молекулы,
Называемые аминокислотами. Амино-кислоты — это и есть те «бусинки», из которых состоит белок, и устроены они сравнительно просто.
КАК УСТРОЕНА АМИНОКИСЛОТА
В каждой молекуле аминокислоты есть атом углерода, связанный с четырьмя заместителями. Один из них — атом водорода, второй — кар-боксильная группа —СООН. Она лег-ко «отпускает на волю» ион водоро-да Н+, благодаря чему в названии аминокислот и присутствует слово «кислота». Третий заместитель — ами-ногруппа — NH 2 и, наконец, четвёр-тый заместитель — группа атомов, ко-торую в общем случае обозначают R . У всех аминокислот R-группы разные, и каждая из них играет свою, очень важную роль.
Свойства «бусинок», отличающие одну аминокислоту от другой, скры-ты в R-группах (их ещё называют бо-ковыми цепями). Что же касается группы — СООН, то химики-органи-ки относятся к ней с большим почте-нием: всем другим атомам углерода в молекуле даются обозначения в зави-симости от степени их удалённости от карбоксильной группы. Ближай-ший к ней атом именуют а-атомом, второй — в-атомом, следующий — у -атомом и т. д. Атом углерода в ами-нокислотах, который находится бли-же всех к карбоксильной группе, т. е. а-атом, связан также с аминогруппой, поэтому природные аминокислоты, входящие в состав белка, называют а-аминокислотами.
В природе встречаются также ами-нокислоты, в которых NH^-группа связана с более отдалёнными от кар-боксильной группы атомами углеро-да. Однако для построения белков природа выбрала именно а-аминокислоты. Это обусловлено прежде всего тем, что только а-аминокислоты, соединённые в длинные цепи, способны обеспечить достаточную прочность и устойчивость структуры больших белковых молекул.
Число а-аминокислот, различа-ющихся R-группой, велико. Но чаще других в белках встречается всего 20 разных аминокислот. Их можно рас-сматривать как алфавит «языка» бел-ковой молекулы. Химики называют эти главные аминокислоты стандарт-ными, основными или нормальными. Условно основные аминокислоты де-лят на четыре класса.
В первый входят аминокислоты с неполярными боковыми цепями. Во второй — аминокислоты, со-держащие полярную группу. Следую-щие два составляют аминокислоты с боковыми цепями, которые могут заряжаться положительно (они объе-диняются в третий класс) или отрица-тельно (четвёртый). Например, диссо-циация карбоксильной группы даёт анион — СОО-, а протонирование ато-ма азота — катион, например — NH 3 + . Боковые цепи аспарагиновой и глута-миновой кислот имеют ещё по одной карбоксильной группе —СООН, кото-рая при значениях рН, характерных для живой клетки (рН = 7), расстаётся с ионом водорода (Н+) и приобрета-ет отрицательный заряд. Боковые це-пи аминокислот лизина, аргинина и гистидина заряжены положительно, поскольку у них есть атомы азота, ко-торые, наоборот, могут ион водорода присоединять.
Каждая а-аминокислота (кроме глицина) в зависимости от взаимно-го расположения четырёх заместите-лей может существовать в двух фор-мах. Они отличаются друг от друга, как предмет от своего зеркального от-ражения или как правая рука от ле-вой. Такие соединения получили название хоральных (от грен. «хир» — «рука»). Хиральные молекулы открыл в 1848 г. великий французский учё-ный Луи Пастер. Два типа оптических изомеров органических молекул по-лучили названия Д-форма (от лат. dexter — «правый») и Z-форма (от лат. laevus — «левый»). Кстати, одно из названий других хиральных моле-кул — глюкозы и фруктозы — декст-роза и левулоза. Примечательно, что в состав белков входят только Z-ами нокислоты, и вся белковая жизнь на Земле — «левая».
Для нормальной жизнедеятельно-сти организм нуждается в полном на-боре из 20 основных a-Z-аминокислот. Но одни из них могут быть синтезиро-ваны в клетках самого организма, а другие — должны поступать в готовом виде из пищевых продуктов. В пер-вом случае аминокислоты называют заменимыми, а во втором — незамени-мыми. Набор последних для разных организмов различен. Например, для белой крысы незаменимыми являют-ся 10 аминокислот, а для молочнокислых бактерий — 16. Растения могут са-мостоятельно синтезировать самые разнообразные аминокислоты, созда-вать такие, которые не встречаются в белках.
Для удобства 20 главных амино-кислот обозначают символами, ис-пользуя одну или первые три буквы русского или английского названия аминокислоты, например аланин — Ала или А, глицин — Гли или G .
ЧТО ТАКОЕ ПЕПТИД
Полимерная молекула белка образует-ся при соединении в длинную цепоч-ку бусинок-аминокислот. Они нани-зываются на нить химических связей благодаря имеющимся у всех амино-кислот амино- и карбоксильной груп-пам, присоединённым к а-атому угле-рода.
Образующиеся в результате такой реакции соединения называются пеп-тидами; (—СО— NH —группировка в них — это пептидная группа, а связь между атомами углерода и азота — пептидная связь (её ещё называют амидной). Соединяя аминокислоты посредством пептидных связей, мож-но получить пептиды, состоящие из остатков очень многих аминокислот. Такие соединения получили название полипептиды. Полипептидное стро-ение белковой молекулы доказал в 1902 г. немецкий химик Эмиль Гер-ман Фишер.
На концах аминокислотной це-почки находятся свободные амино-и карбоксильная группы; эти концы цепочки называют N - и С-концами. Аминокислотные остатки в полипеп-тидной цепочке принято нумеровать с N-конца.
Общее число аминокислотных ос-татков в белковой молекуле изменя-ется в очень широких пределах. Так, человеческий инсулин состоит из 51 аминокислотного остатка, а лизо-цим молока кормящей матери — из 130. В гемоглобине человека 4 ами-нокислотные цепочки, каждая из которых построена из примерно 140 аминокислот. Существуют белки, имеющие почти 3 тыс. аминокис-лотных остатков в единой цепи.
Молекулярные массы белков лежат в диапазоне примерно от 11 тыс. для малых белков, состоящих из 100 ами-нокислотных остатков, до 1 млн и бо-лее для белков с очень длинными полипептидными цепями или для белков, состоящих из нескольких по-липептидных цепей.
Возникает вопрос: как же всё ог-ромное многообразие белков с раз-личными функциями и свойствами может быть создано всего из 20 мо-лекул? А разгадка этого секрета при-роды проста — каждый белок имеет свой неповторимый аминокислот-ный состав и уникальный порядок со-единения аминокислот, называемый первичной структурой белка.
СПИРАЛИ И СЛОИ
В начале 50-х гг. XX в. американские химики Лайнус Карл Полинг (1901— 1994), награждённый Нобелевской премией за исследования природы химической связи, и Роберт Кори (1897—1971) предположили, что не-которые участки аминокислотной це-почки в белках закручены в спираль. Благодаря совершенствованию экс-периментальных методов (структуру белков изучают с помощью рентгенов-ских лучей) через несколько лет эта гениальная догадка подтвердилась.
Действительно, полипептидные цепи очень часто образуют спираль, закрученную в правую сторону. Это первый, самый низкий уровень про-странственной организации белко-вых цепочек Здесь-то и начинают иг-рать роль слабые взаимодействия «бусинок»-аминокислот: группа С=0 и группа N — H из разных пептидных связей могут образовывать между со-бой водородную связь. Оказалось, что в открытой Полингом и Кори спирали такая связь образована меж-ду группой С=0 каждой г-й аминокис-лоты и группой N — H (i + 4)-й амино-кислоты, т. е. между собой связаны аминокислотные остатки, отстоящие друг от друга на четыре «бусинки». Эти водородные связи и стабилизиру-ют такую спираль в целом. Она полу-чила название a.-спирали.
Позднее выснилось, что а-спираль — не единственный способ ук-ладки аминокислотных цепочек. По-мимо спиралей они образуют ещё и слои. Благодаря всё тем же водород-ным связям между группами С=0 и N — H друг с другом могут «слипаться» сразу несколько разных фрагментов одной полипептидной цепи. В резуль-тате получается целый слой — его на-звали ^-слоем.
В большинстве белков а-спирали и р-слои перемежаются всевозможными изгибами и фрагментами цепи без какой-либо определённой структуры. Когда имеют дело с пространствен-ной структурой отдельных участков белка, говорят о вторичной структу-ре белковой молекулы.
БЕЛОК В ПРОСТРАНСТВЕ
Для того чтобы получить полный «портрет» молекулы белка, знания первичной и вторичной структуры недостаточно. Эти сведения ещё не дают представления ни об объёме, ни о форме молекулы, ни тем более о расположении участков цепи по отношению друг к другу. А ведь все спирали и слои каким-то образом размещены в пространстве. Общая пространственная структура поли-пептидной цепи называется третич-ной структурой белка.
Первые пространственные модели молекул белка — миоглобина и гемо-глобина — построили в конце 50-х гг. XX в. английские биохимики Джон Ко-удери Кендрю (родился в 1917 г.) и Макс Фердинанд Перуц (родился в 1914 г.). При этом они использовали данные экспериментов с рентгенов-скими лучами. За исследования в об-ласти строения белков Кендрю и Перуц в 1962 г. были удостоены Нобе-левской премии. А в конце столетия была определена третичная структура уже нескольких тысяч белков.
При образовании третичной струк-туры белка наконец-то проявляют активность R-группы — боковые це-пи аминокислот. Именно благодаря им «слипаются» между собой боль-шинство «бусинок»-аминокислот, придавая цепи определённую форму в пространстве.
В живом организме белки всегда находятся в водной среде. А самое большое число основных аминокис-лот — восемь — содержат неполяр-ные R-группы. Разумеется, белок стремится надёжно спрятать внутрь своей молекулы неполярные боковые цепи, чтобы ограничить их контакт с водой. Учёные называют это воз-никновением гидрофобных взаимо-действий (см. статью «Мельчайшая единица живого»).
Благодаря гидрофобным взаимо-действиям вся полипептидная цепоч-ка принимает определённую форму в пространстве, т. е. образует третич-ную структуру.
В молекуле белка действуют и дру-гие силы. Часть боковых цепей основ-ных аминокислот заряжена отрица-тельно, а часть — положительно. Так как отрицательные заряды притяги-ваются к положительным, соответст-вующие «бусинки» «слипаются». Элек-тростатические взаимодействия, или, как их называют иначе, солевые мос-тики, — ещё одна важная сила, ста-билизирующая третичную структуру.
У семи основных аминокислот есть полярные боковые цепи. Между ними могут возникать водородные связи, тоже играющие немалую роль в поддержании пространственной структуры белка.
Между двумя аминокислотными остатками цистеина иногда образу-ются ковалентные связи (— S —S—), которые очень прочно фиксируют расположение разных участков бел-ковой цепи по отношению друг к другу. Такие связи называют дисуль-фидными мостиками. Это самые не-многочисленные взаимодействия в белках (в некоторых случаях они во-обще отсутствуют), зато по прочно-сти они не имеют равных.
ВЫСШИЙ УРОВЕНЬ ПРОСТРАНСТВЕННОЙ ОРГАНИЗАЦИИ БЕЛКОВ
Молекула белка может состоять не из одной, а из нескольких полипептидных цепей. Каждая такая цепь представляет собой самостоятельную пространственную структуру — субь-единицу. Например, белок гемогло-бин состоит из четырёх субъединиц, которые образуют единую молекулу, располагаясь в вершинах почти пра-вильного тетраэдра. Субъединицы «прилипают» друг к другу благодаря тем же самым силам, что стабилизи-руют третичную структуру. Это гид-рофобные взаимодействия, солевые мостики и водородные связи.
Если белок состоит из нескольких субъединиц, говорят, что он обладает четвертичной структурой. Такая структура представляет собой высший уровень организации белковой моле-кулы. В отличие от первых трёх уров-ней четвертичная структура есть дале-ко не у всех белков. Приблизительно половина из известных на сегодняш-ний день белков её не имеют.
ПОЧЕМУ БЕЛКИ БОЯТСЯ ТЕПЛА
Связи, поддерживающие пространст-венную структуру белка, довольно лег-ко разрушаются. Мы с детства знаем, что при варке яиц прозрачный яич-ный белок превращается в упругую белую массу, а молоко при скисании загустевает. Происходит это из-за раз-рушения пространственной структуры белков альбумина в яичном белке и ка-зеина (огглат. caseus — «сыр») в моло-ке. Такой процесс называется денату-рацией. В первом случае её вызывает нагревание, а во втором — значи-тельное увеличение кислотности (в результате жизнедеятельности обита-ющих в молоке бактерий). При дена-турации белок теряет способность выполнять присущие ему в организме функции (отсюда и название процес-са: от лат. denaturare — «лишать при-родных свойств»). Денатурированные белки легче усваиваются организмом, поэтому одной из целей термической обработки пищевых продуктов яв-ляется денатурация белков.
ЗАЧЕМ НУЖНА ПРОСТРАНСТВЕННАЯ СТРУКТУРА
В природе почти ничего не происхо-дит случайно. Если белок принял определённую форму в пространстве, это должно служить достижению ка-кой-то цели. Действительно, только бе-лок с «правильной» пространственной структурой может обладать опреде-лёнными свойствами, т. е. выполнять те функции в организме, которые ему предписаны. А делает он это с помо-щью всё тех же R-групп аминокислот. Оказывается, боковые цепи не толь-ко поддерживают «правильную» фор-му молекулы белка в пространстве. R-группы могут связывать другие орга-нические и неорганические молекулы, принимать участие в химических ре-акциях, выступая, например, в роли ка-тализатора.
Часто сама пространственная ор-ганизация полипептидной цепи как раз" и нужна для того, чтобы сосредо-точить в определённых точках про-странства необходимый для выполне-ния той или иной функции набор боковых цепей. Пожалуй, ни один процесс в живом организме не прохо-дит без участия белков.
В ЧЁМ СЕКРЕТ ФЕРМЕНТОВ
Все химические реакции, протекаю-щие в клетке, происходят благодаря особому классу белков — фермен-там. Это белки-катализаторы. У них есть свой секрет, который позволяет им работать гораздо эффективнее других катализаторов, ускоряя реак-ции в миллиарды раз.
Предположим, что несколько при-ятелей никак не могут встретиться. Но стоило одному из них пригласить друзей на день рождения, как резуль-тат не заставил себя ждать: все оказа-лись в одном месте в назначенное время.
Чтобы встреча состоялась, понадо-билось подтолкнуть друзей к контак-ту. То же самое делает и фермент. В его молекуле есть так называемые центры связывания. В них расположе-ны привлекательные для определён-ного типа химических соединений (и только для них!) «уютные кресла» — R-группы, связывающие какие-то уча-стки молекул реагирующих веществ. Например, если одна из молекул име-ет неполярную группу, в центре свя-зывания находятся гидрофобные бо-ковые цепи. Если же в молекуле есть отрицательный заряд, его будет под-жидать в молекуле фермента R-груп па с положительным зарядом.
В результате обе молекулы реаген-тов связываются с ферментом и ока-зываются в непосредственной близо-сти друг от друга. Мало того, те их группы, которые должны вступить в химическую реакцию, сориентирова-ны в пространстве нужным для реак-ции образом. Теперь за дело прини-маются боковые цепи фермента, играющие роль катализаторов. В фер-менте все «продумано» таким обра-зом, что R-группы-катализаторы тоже расположены вблизи от места собы-тий, которое называют активным центром. А после завершения реак-ции фермент «отпускает на волю» мо-лекулы-продукты (см. статью «Фер-менты — на все руки мастера»).
ОТКУДА БЕРЁТСЯ ИММУНИТЕТ
Белки выполняют в организме мно-жество функций; они, например, за-щищают клетки от нежелательных вторжений, предохраняют их от по-вреждений. Специальные белки — антитела обладают способностью распознавать проникшие в клетки бактерии, вирусы, чужеродные поли-мерные молекулы и нейтрализовывать их.
У высших позвоночных от чуже-родных частиц организм защищает иммунная система. Она устроена так, что организм, в который вторг-лись такие «агрессоры» — антигены, начинает вырабатывать антитела. Молекула антитела прочно связыва-ется с антигеном: у антител, как и у ферментов, тоже есть центры связы-вания. Боковые цепи аминокислот расположены в центрах таким обра-зом, что антиген, попавший в эту ло-вушку, уже не сможет вырваться из «железных лап» антитела. После свя-зывания с антителом враг выдворяет-ся за пределы организма.
Можно ввести в организм неболь-шое количество некоторых полимер-ных молекул, входящих в состав бак-терий или вирусов-возбудителей какой-либо инфекционной болезни.
В организме немедленно появятся соответствующие антитела. Теперь попавший в кровь или лимфу «насто-ящий» болезнетворный микроб тот-час же подвергнется атаке этих анти-тел, и болезнь будет побеждена. Такой способ борьбы с инфекцией есть не что иное, как нелюбимая многими прививка. Благодаря ей организм приобретает иммунитет к инфекци-онным болезням.
ДЛЯ ЧЕГО В ГЕМОГЛОБИНЕ ЖЕЛЕЗО
В природе существуют белки, в ко-торых помимо аминокислот содер-жатся другие химические компонен-ты, такие, как липиды, сахара, ионы металлов. Обычно эти компоненты играют важную роль при выполне-нии белком его биологической функ-ции. Так, перенос молекул и ионов из одного органа в другой осуществля-ют транспортные белки плазмы крови. Белок гемоглобин (от греч. «гема» — «кровь» и лат. globus — «шар», «шарик»), содержащийся в кровяных клетках — эритроцитах (от греч. «эритрос» — «красный» и «китос» — «клетка»), доставляет кис-лород от лёгких к тканям. В молеку-ле гемоглобина есть комплекс иона железа Fe 24 " со сложной органической молекулой, называемый гемам. Гемо-глобин состоит из четырёх белковых субъединиц, и каждая из них содер-жит по одному гему.
В связывании кислорода в лёгких принимает участие непосредственно ион железа. Как только к нему хотя бы в одной из субъединиц присоединя-ется кислород, сам ион тут же чуть-чуть меняет своё расположение в мо-лекуле белка. Движение железа «про-воцирует» движение всей аминокис-лотной цепочки данной субъединицы, которая слегка трансформирует свою третичную структуру.
Другая субъеди-ница, ещё не присоединившая кислород, «чувствует», что произошло с со-седкой. Её структура тоже начинает меняться. В итоге вторая субъедини-ца связывает кислород легче, чем пер-вая. Присоединение кислорода к третьей и четвёртой субъединицам происходит с ещё меньшими трудно-стями. Как видно, субъединицы помо-гают друг другу в работе. Для этого-то гемоглобину и нужна четвертичная структура. Оксид углерода СО (в про-сторечии угарный газ) связывается с железом в геме в сотни раз прочнее кислорода. Угарный газ смертельно опасен для человека, поскольку ли-шает гемоглобин возможности при-соединять кислород.
А ЕЩЁ БЕЛКИ...
Служат питательными веществами. В семенах многих растений (пшени-цы, кукурузы, риса и др.) содержатся пищевые белки. К ним относятся так-же альбумин — основной компонент яичного белка и казеин — главный белок молока. При переваривании в организме человека белковой пищи происходит гидролиз пептидных свя-зей. Белки «разбираются» на отдель-ные аминокислоты, из которых орга-низм в дальнейшем «строит» новые пептиды или использует для полу-чения энергии. Отсюда и название:
Греческое слово «пептос» означает «переваренный». Интересно, что гид-ролизом пептидной связи управляют тоже белки — ферменты.
Участвуют в регуляции клеточ-ной и физиологической активности. К подобным белкам относятся мно-гие гормоны (от греч. «гормао» — «по-буждаю»), такие, как инсулин, регули-рующий обмен глюкозы, и гормон роста.
Наделяют организм способно-стью изменять форму и передвигать-ся. За это отвечают белки актин и ми-озин, из которых построены мышцы.
Выполняют опорную и защитную функции, скрепляя биологические структуры и придавая им прочность. Кожа представляет собой почти чис-тый белок коллаген, а волосы, ногти и перья состоят из прочного нерас-творимого белка кератина.
ЧТО ЗАПИСАНО В ГЕНАХ
Последовательность аминокислот в белках кодируется генами, которые хранятся и передаются по наследству с помощью молекул ДНК (см. статьи «Хранитель наследственной инфор-мации. ДНК» и «Экспрессия генов»). Пространственную структуру белка задаёт именно порядок расположе-ния аминокислот. Получается, что не только первичная, но и вторичная, третичная и четвертичная структуры белков составляют содержание на-следственной информации. Следо-вательно, и выполняемые белками функции запрограммированы гене-тически. Громадный перечень этих функций позволяет белкам по праву называться главными молекулами жизни. Поэтому сведения о белках и есть то бесценное сокровище, кото-рое передаётся в природе от поколе-ния к поколению.
Интерес человека к этим органи-ческим соединениям с каждым годом только увеличивается. Сегодня учёные уже расшифровали структуру многих белковых молекул. Они выясняют функции самых разных белков, пыта-ются определить взаимосвязь функ-ций со структурой. Установление сходства и различий у белков, выпол-няющих аналогичные функции у раз-ных живых организмов, позволяет глубже проникать в тайны эволюции.
АМИНОКИСЛОТЫ — ПОКАЗАТЕЛИ ВОЗРАСТА
D - и L -формы аминокислот обладают способностью очень медленно превращаться друг в друга. За определённый (весьма длительный) период времени чистая D- или I-форма может стать смесью равных количеств обеих форм. Такая смесь называется раиемагом, а сам процесс —раие-мизаиией. Скорость рацемизации зависит от температуры и типа амино-кислоты. Данное свойство можно использовать для определения возрас-та ископаемых остатков организмов, а при необходимости — и живых существ. Например, в белке дентина (дентин — костная ткань зубов) 1-ас-парагиновая кислота самопроизвольно раиемизуется со скоростью 0,1 % в год. У детей в период формирования зубов в дентине содержится толь-ко 1-аспарагиновая кислота. Дентин выделяют из зуба и определяют В нём содержание 0-формы. Результаты теста достаточно точны. Так, для 97-лет-ней женщины, возраст которой был документально засвидетельствован, тест показал возраст 99 лет. Данные исследований, выполненных на ис-копаемых остатках доисторических животных — слонов, дельфинов, мед-ведей, — хорошо согласуются с результатами датирования, полученными радионуклидным методом.
ЗА ЧТО СЕНГЕР ПОЛУЧИЛ НОБЕЛЕВСКИЕ ПРЕМИИ
При гидролизе белков до аминокислот (разрушении пептидной связи во-дой) теряется информация о последовательности их соединения. Поэто-му долгое время считали, что определение первичной структуры белка представляет собой совершенно безнадежную задачу. Но в 50-х гг. XX в. английский биохимик Фредерик Сенгер (родился в 1918 г.) смог расшиф-ровать последовательность аминокислот в полипептидных цепях гормо-на инсулина. За эту работу, на выполнение которой ушло несколько лет, в 1958 г. Сенгер был удостоен Нобелевской премии по химии (двадца-тью годами позже он совместно с У. Гилбертом получил вторую премию за вклад в установление первичной структуры ДНК).
Принципы определения аминокислотной последовательности, впервые сформулированные Сенгером, используются и ныне, правда, со всевоз-можными вариациями и усовершенствованиями. Процедура установле-ния первичной структуры белка сложна и многоступенчата: в ней около десятка различных стадий. Сначала белок расщепляют до отдельных ами-нокислот и устанавливают их тип и количество в данном веществе. На сле-дующей стадии длинную белковую молекулу расщепляют уже не полно-стью, а на фрагменты. Затем в этих фрагментах определяют порядок соединения аминокислот, последовательно отделяя их одну за другой. Расшепление белка на фрагменты проводят несколькими способами, что-бы в разных фрагментах были перекрывающиеся участки. Выяснив поря-док расположения аминокислот во всех фрагментах, получают полную ин-формацию о том, как аминокислоты расположены в белке. К концу XX в. созданы специальные приборы, определяющие последовательность амино-кислот в молекуле белка в автоматическом режиме — секвенаторы (от англ. sequence — «последовательность»).
МОЛОКО И КИСЛОМОЛОЧНЫЕ ПРОДУКТЫ
Молоко представляет собой коллоидный раствор жира в воде. Под микроскопом хорошо видно, что оно неоднородно: в бесцветном растворе (сыворотке) плавают жировые шарики.
В коровьем молоке обычно содержится от 3 до 6 % жиров (в основном это сложные эфиры глицерина и насыщенных карбоновых кислот - пальмитиновой, стеариновой), около 3 % белков, а ешё углеводы, органические кислоты, витамины и минеральные вещества.
Белок казеин в молоке присутствует в связанном виде - ковалентно присоединённые к аминокислоте сери-ну фосфатные группы образуют соли с ионами кальция. При подкислении молока эти соли разрушаются, и казеин выделяется в виде белой творожистой массы. В желудке человека под действием особых ферментов происходит процесс, называемый “створажива-нием казеина”. Створоженный казеин выпадает в осадок и медленнее выводится из организма, а потому полнее усваивается. Казеин высоко питателен:
В нём есть почти все аминокислоты, необходимые человеку для построения собственных белков. В чистом виде он представляет собой безвкусный белый порошок, не растворимый в воде. Помимо него в молоке содержатся и другие белки, например лактальбумин. При кипячении этот белок превращается в нерастворимую форму, образуя на поверхности кипячёного молока характерную белую плёнку - пенку.
Входящий в состав молока сахар лактоза С^НддО, изомерен сахарозе. В организме человека под действием фермента лактазы этот сахар расщепляется на моносахариды глюкозу и галактозу, которые легко усваиваются. За счёт этого, например, грудные дети пополняют запасы углеводов. Интересно, что у многих людей (в основном у представителей монголоидной расы) организм в зрелом возрасте утрачивает способность расщеплять лактозу.
Проходя через пищеварительный тракт, лактоза не усваивается, а становится питательной средой для развития различных болезнетворных микроорганизмов, что приводит к общему недомоганию. Именно поэтому народы Дальнего Востока (японцы, китайцы) практически не употребляют в пишу молочные продукты.
В промышленных условиях молоко подвергают тепловой обработке, цель которой - подавить развитие микроорганизмов и продлить срок его хранения. Для этого молоко пастеризуют - выдерживают 30 мин при 65 °С, а также используют кратковременную термообработку - нагревают в течение 10-20 с до 71 °С. По сравнению с пастеризацией термообработка лучше сохраняет питательные вещества, в первую очередь витамины. Чтобы молоко не расслаивалось на сливки и сыворотку, его гомогенизируют - пропускают под давлением через небольшие отверстия. Жировые шарики дробятся, уменьшаются в размерах, а молоко становится более вязким.
Значительная часть молока идёт на переработку - для производства сливочного масла, сыра и кисломолочных продуктов (кефира, ряженки, простокваши, сметаны).
Чтобы получить кефир, молоко сквашивают - выдерживают в течение 8-10 ч при 20-25 °С, добавляя затравку молочнокислых бактерий. Под их действием лактоза распадается до молочной кислоты:
С„н„о„ + н,о =лактоза == 4СНзСН(ОН)СООН. молочная (2-гидроксипропановая) кислота
Именно молочная кислота определяет специфический вкус кефира. По мере того как она накапливается в растворе, происходит коагуляция (свёртывание)казеина, который выделяется в свободном виде. Поэтому кефир имеет более густую консистенцию, чем молоко. Молочнокислое сбраживание лактозы сопровождается спиртовым брожением, из-за чего в кисломолочных продуктах, в частности в кефире, есть небольшое количество алкоголя (до 0,03 %). В кисломолочных продуктах содержатся также микроорганизмы, которые подавляют развитие болезнетворных бактерий и тем самым улучшают пишеварение.
Творог тоже получают сквашиванием молока молочнокислыми бактериями. Его главной составной частью является белок казеин.
Чтобы приготовить сливочное масло, от молочной сыворотки необходимо отделить капельки жира, входящие в состав молока. Для этого сбивают сливки - верхний, более жирный слой, образующийся при отстаивании молока.
Казеин входит также в состав сыров. Их делают, добавляя в молоко бактериальную закваску и специальные ферменты, а затем подогревая смесь до определённой температуры. В выделившийся сгусток вновь вводят ферменты и подогревают. При этом происходит частичное изменение структуры и состава казеина. Затем смесь раскладывают по формам и длительное время - до шести месяцев - выдерживают при низкой температуре (не выше 15 °С). Во время созревания казеин под действием ферментов распадается на поли-пептиды и свободные аминокислоты. Часть аминокислот окисляется кислородом воздуха, при этом образуются аммиак, альдегиды, а также кетокислоты, придающие сыру характерный аромат.
Скисание молока - привычный пример денатурации белка.
МЕДНАЯ КРОВЬ
В холодных водах Перуанского течения в Тихом океане обитает кальмар Dosidicus gigas. Его сигарообразное тело вместе со щупальцами достигает в длину 3,5 м, а масса гиганта может превышать 150 кг. Мощные мышиы выбрасывают струю воды с силой, с какой она бьёт из пожарного рукава, благодаря чему кальмар способен двигаться со скоростью до 40 км/ч. Клювом, очень крепким и острым, он может перебить стальной кабель. По свидетельству очевидцев, кальмар буквально в клочья раздирает 20-килограммовую рыбину. Этот свирепый хишник очень опасен и для человека. В книге Франка Лейна “Царство осьминога” утверждается, что “человек, упавший за борт в местах, где обитает много кальмаров, не проживёт и полминуты”.
Чтобы “зарядиться” энергией, этому обитателю океана требуется много кислорода - не менее 50 л в час. По-ступаюший из морской воды кислород разносится по телу кальмара с помошью особого белка, содержащего медь, - гемоиианина (от греч. “гема” - “кровь” и “кианос” - “лазурный”, “голубой”).
Стоит заметить, что в крови позвоночных кислород “транспортируют” атомы железа в составе гема - особой сложной молекулы, которая входит в состав белка гемоглобина. Им буквально нашпигованы красные кровяные клетки - эритроциты. Молекула гемоглобина содержит четыре гемовых фрагмента, каждый из которых способен связать молекулу кислорода. В отличие от гемоглобина, в гемоиианине атомы меди непосредственно связаны с белковыми молекулами, которые не включены ни в какие клетки, а свободно “плавают” в крови. Зато одна молекула гемоииани
На способна связать до 200 атомов меди. И ешё одна особенность гемоииани-на - его молекулы имеют огромные даже для белков размеры. У “обычных” белков, входящих в состав яиц, молока, мыши, молекулярная масса колеблется в пределах от б тыс. до 1 млн, а молекулярная масса гемоиианина может достигать 10 млн! Это один из самых крупных белков; больше по размеру и массе только белковые комплексы у вирусов.
Гемоиианин - очень древний белок. Он устроен проще, чем гемоглобин и не так эффективен. Тем не менее при малом содержании кислорода в морской воде гемоиианин довольно успешно снабжает им ткани холоднокровных животных. Так, давление кислорода в жабрах лангуста составляет всего 7 мм рт. ст. (930 Па), а в тканях - 3 мм рт. ст.; причём концентрация этого газа в крови лангуста в 20 раз выше, чем в морской воде.
Кроме кальмаров, кислород переносится “голубой кровью” также у де-сятиногих ракообразных (омары, крабы, креветки). Гемоиианин найден у всех головоногих моллюсков (осьминоги, кальмары, каракатицы), разнообразных улиток, пауков и др. А вот у морских гребешков, устриц и других двустворчатых моллюсков его нет.
Количество гемоиианина в крови может быть самым разным. Так, у шустрых осьминога и мечехвоста (морское животное типа членистоногих) концентрация этого необычного белка доходит до 10 г в 100 мл крови - почти столько же гемоглобина в крови человека. В то же время, у малоподвижного съедобного моллюска морское ушко Hatiotis tuberculata в 100 мл крови всего 0,03 г гемоиианина. Это и понятно: чем более активно животное,
Чем больше кислорода необходимо ему для восполнения энергетических затрат, тем выше в крови концентрация белка, переносящего кислород.
Гемоиианин был открыт в 60-х гг. XIX в., когда биологи заметили, что кровь головоногих моллюсков при прохождении через жабры окрашивается в голубой цвет. А в 1878 г. бельгийский физиолог Леон Фредерик доказал, что голубой цвет вызван реакцией кислорода с медьсодержащим белком, который он назвал гемоиианином. Когда последний теряет кислород, он, в отличие от гемоглобина, становится бесцветным. Примечательно, что всю работу по изучению нового белка Фредерик выполнил в течение одного дня.
Из гемоиианина нетрудно полностью извлечь медь. Аля этого достаточно обработать белок в отсутствие кислорода реактивом, который прочно связывается с ионами одновалентной меди. Таким же способом можно определить содержание меди в гемоиианине. Лишённый этого металла, он теряет способность переносить кислород. Но если потом ввести в раствор белка ионы Си"1", гемоиианин восстанавливает свою физиологическую активность.
Так было доказано, что в отсутствие кислорода медь гемоиианина находится в степени окисления +1. При избытке же этого газа происходит частичное окисление металла. При этом всегда на одну связанную гемоиианином молекулу кислорода приходится два атома меди. Таким образом, кислород окисляет ровно половину атомов меди. Это ещё одно отличие гемоиианина от значительно более распространённого в животном мире гемоглобина, в котором все атомы железа равноценны и имеют заряд +2 как в свободном состоянии, так и в комплексе с кислородом.
Среди органических веществ белки , или протеины , - самые многочисленные, наиболее разнообразные и имеющие первостепенное значение биополимеры. На их долю приходится 50 - 80% сухой массы клетки.
Молекулы белков имеют большие размеры, поэтому их называют макромолекулами . Кроме углерода , кислорода , водорода и азота , в состав белков могут входить сера, фосфор и железо. Белки отличаются друг от друга числом (от ста до нескольких тысяч), составом и последовательностью мономеров. Мономерами белков являются аминокислоты (рис. 1)
Бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Каждая аминокислота имеет свое название, особое строение и свойства. Их общую формулу можно представить в следующем виде:

Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (-NH 2 ) с основными свойствами, другая - карбоксильной группой (-COOH ) с кислотными свойствами. Часть молекулы, называемая радикалом (R ), у разных аминокислот имеет различное строение. Наличие в одной молекуле аминокислоты основной и кислотной групп обусловливает их высокую реакционную способность. через эти группы происходит соединение аминокислот при образовании белка. При этом возникает молекула воды, а освободившиеся электроны образуют пептидную связь. Поэтому белки называют полипептидами .
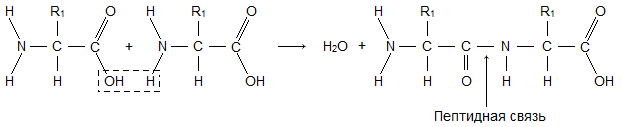
Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
Последовательность аминокислот в составе полипептидной цепи представляет первичную структуру
белка. Она уникальна для любого белка и определяет его форму, свойства и функции.
Большинство белков имеют вид спирали в результате образования водородных связей между -CO-
и -NH-
группами разных аминокислотных остатков полипептидной цепи. Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль - вторичная структура
белка.
Третичная структура - трехмерная пространственная «упаковка» полипептидной цепи. В результате возникает причудливая, но для каждого белка специфическая конфигурация - глобула . Прочность третичной структуры обеспечивается разнообразными связями, возникающими между радикалами аминокислот.
Четвертичная структура
характерна не для всех белков. Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс. Например, гемоглобин крови человека представляет комплекс из четырех макромолекул белка.
Такая сложность структуры белковых молекул связана с разнообразием функций, свойственных этим биополимерам.
Нарушение природной структуры белка называют денатурацией
. Она может происходить под воздействием температуры, химических веществ, лучистой энергии и других факторов. При слабом воздействии распадается только четвертичная структура, при более сильном - третичная, а затем - вторичная, и белок остается в виде полипептидной цепи.
Этот процесс частично обратим: если не нарушена первичная структура, то денатурированный белок способен восстанавливать свою структуру. Отсюда следует, что все особенность строение макромолекулы белка определяются его первичной структурой.
Кроме простых белков , состоящих только из аминокислот, есть еще и сложные белки